Будова и функции на РНК. транспортна РНК. Будова, функции. Схема на молекулата на тРНК
Є синтез на протеинова молекула от уран на матрична РНК (транслация). Въпреки това, по отношение на транскрипцията, нуклеотидната последователност не може да бъде директно преведена в аминокиселина; Следователно, за транслацията на транслацията е необходим посредник в подобна транспортна РНК (тРНК), чиято функция е транслацията на генетичния код към „мова” на аминокиселините.
Глобална характеристика на трансферната РНК
Транспортната РНК или тРНК са малки молекули, които доставят аминокиселини до мястото на протеинов синтез (рибозоми). Броят на този тип рибонуклеинова киселина в клитините става приблизително 10% от общия пул РНК.
Подобно на други разновидности, тРНК се образува от рибонуклеозид трифосфатно копие. Дължината на нуклеотидната последователност е 70-90 ленти и около 10% от молекулярната структура съдържа второстепенни компоненти.
Чрез тези, които кожните аминокиселини образуват своя носител като тРНК, клитинът синтезира голям брой различни молекули. Угар под формата на жив организъм, индикаторът му варира от 80 до 100.
Функции на тРНК
Транспортната РНК е пост-главен субстрат за протеиновия синтез, който се намира в рибозомите. Поради уникалността на връзката с аминокиселините и с матричната последователност на tRNA, функцията на сензорния адаптер се транспонира под формата на протеин. Взаимодействието на такъв посредник с матрица, която кодира, подобно на транскрипцията, се основава на принципа на комплементарност на азотните бази.
Главната функция на tRNA участва в приемането на аминокиселинните фенери и последващия им транспорт до апарата за синтез на протеини. Зад този технически процес стои едно голямо биологично усещане – внедряването на генетичния код. Развитието на този процес се основава на обидни характеристики:
- uxi аминокиселините са кодирани от триплети нуклеотиди;
- за кожен триплет (или кодон) има антикодон, който влиза преди склада на tRNA;
- дермалната тРНК може да се свърже само с пееща аминокиселина.

По този начин аминокиселинната последователност на протеина се определя от факта, че тРНК в този ред ще бъде комплементарна на взаимодействието с информационната РНК в процеса на транслация. Възможно е да има признаци на проявление в транспортната РНК на функционални центрове, единият от които се дължи на добавянето на аминокиселина, а другият се дължи на свързване с кодон. Следователно функциите са тясно свързани.
Будова транспортна РНК
Уникалността на tRNA е, че нейната молекулярна структура не е линейна. Вон включва спирални dvolantsyugkovy летви, които се наричат стъбла, и 3 odnolyantsyug бримки. Зад формата, такава конформация отгатва стабилния лист.
Структурата на tRNA има следните стебла:
- акцептор;
- антикодон;
- дихидроуридил;
- псевдоуридилий;
- допълнителен.
Спираловидни стъбла под виното могат да бъдат отстранени от 5 до 7 двойки Watson-Crickson. В края на акцепторното стъбло на гниене има малко копие от несдвоени нуклеотиди, 3-хидроксил, което е деформирана молекула на аминокиселина.

Структурната област за свързване на иРНК е една от бримките на тРНК. Възможно е да се отмъсти на антикодона, който е комплементарен към триплета на значението на самия антикодон и приемащия край, за да се осигури адапторната функция на тРНК.
Третинна структура на молекулата
„Стабилният лист“ е вторичната структура на тРНК, протео за сгъване на молекулата приема L-подобна конформация, сякаш е свързана с допълнителни водни връзки.
L-формата има третичната структура на tRNA и се състои от две почти перпендикулярни A-RNA спирали, които са дълги 7 nm и дълги 2 nm. Тази форма на молекулата е по-малка от 2 цикъла, единият от които съдържа антикодон, а другият има акцепторен център.

Особености на свързването на тРНК с аминокиселина
Активирането на аминокиселини (добавяне към транспортната РНК) се медиира от аминоацил-тРНК синтетаза. Този ензим изпълнява едновременно 2 важни функции:
- катализира установяването на ковалентна връзка между 3`-хидроксилната група на акцепторното стебло и аминокиселината;
- гарантират принципа на избор на жизнеспособност.
За кожата те имат своя собствена аминоацил-тРНК синтетаза. Той може да взаимодейства само с различен тип транспортна молекула. Tse означава, че оставащият антикодон може да бъде комплементарен на триплета, който кодира същата аминокиселина. Например, синтетазата на левцин се инхибира само по отношение на тРНК за левцин.
Молекулата на аминоацил-тРНК синтетазата има три нуклеотид-свързващи чревни конформации и заряди, които са комплементарни на нуклеотидите на двойния антикодон в тРНК. По този начин ензимът обозначава необходимата транспортна молекула. Обогатена с фрагмент на разпознаване е нуклеотидната последователност на акцепторното стебло.
РНК- полимер, мономери като напр рибонуклеотиди. На повърхността на ДНК РНК не се състои от два, а от един полинуклеотиден ланцет (виняток - деак РНК вирусите могат да удвоят РНК). Нуклеотидите на РНК са способни да установяват водни връзки помежду си. РНК копията са значително по-къси от ДНК копията.
РНК мономер - нуклеотид (рибонуклеотид)- съставен е от излишъци на три вещества: 1) азотна основа, 2) поликарбонатен монозахарид (пентоза) и 3) фосфорна киселина. Азотните основи на РНК също се класифицират като пиримидини и пурини.
Пиримидиновите бази на РНК са урацил, цитозин, пуриновите бази са аденин и гуанин. Монозахаридният РНК нуклеотид е представен от рибоза.
виж три вида РНК: 1) информационен(матрична) РНК – iRNA (mRNA), 2) транспортРНК - тРНК, 3) рибозомаРНК - рРНК.
Всички видове РНК с неразградими полинуклеотиди могат да имат специфична пространствена конформация и да участват в процесите на протеинов синтез. В ДНК се съхранява информация за съществуването на всички видове РНК. Процесът на синтез на РНК върху ДНК матрица се нарича транскрипция.

Транспортна РНКизберете 76 (за 75 до 95) нуклеотиди; молекулно тегло - 25 000-30 000. На част от тРНК се пада приблизително 10% от общата РНК в клитина. Функции на тРНК: 1) транспорт на аминокиселини до мястото на протеинов синтез, рибозоми, 2) транслационен медиатор. Близо 40 вида тРНК се намират в клитините и кожата от тях може да се характеризира с по-малка последователност от нуклеотиди. Във всички тРНК обаче има цаца от вътрешномолекулни комплементарни клетки, чрез които тРНК образуват конформации, които отгатват формата на стабилен лист. Дали тРНК прави примка за контакт с рибозомата (1), антикодонна примка (2), примка за контакт с ензима (3), акцепторен ствол (4), антикодон (5). Аминокиселината се свързва до 3" края на акцепторното стебло. Антикодон- Три нуклеотида, които "разпознават" кодона на iRNA. От това следва, че определена tRNA може да транспортира една аминокиселина, което показва антикодон. Специфичността на половината аминокиселина и тРНК достига границата на мощността на ензима аминоацил-тРНК синтетаза.
Рибозомна РНКпочистване на 3000-5000 нуклеотида; молекулно тегло - 1000000-1500000. Част от рРНК представлява 80-85% от общото количество РНК в клитина. В комплекса с рибозомните протеини рРНК съдържа рибозоми - органоиди, които индуцират протеиновия синтез. В еукариотните клетки синтезът на рРНК се извършва в ядрата. Функции на рРНК: 1) необходимият структурен компонент на рибозомата, като по този начин осигурява функционирането на рибозомата; 2) осигуряване на взаимодействието на рибозомата и тРНК; 3) кочанна връзка на рибозома и кодон-инициатор на iRNA и обозначаване на четящата рамка; 4) формоване на активния център на рибозомата.
Нуклеиновите киселини са реч с високо молекулно тегло, които са съставени от мононуклеотиди, които са свързани един по един в полимерни копия с помощта на 3,5"-фосфодиестерни връзки и са опаковани в зацепки в определен ред.
Нуклеиновите киселини са биополимери от два различни типа: рибонуклеинова киселина (РНК) и дезоксирибонуклеинова киселина (ДНК). Кожният биополимер се състои от нуклеотиди, които се диференцират от въглехидратния излишък (рибоза, дезоксирибоза) и една от азотните бази (урацил, тимин). Vydpovidno на tsikh vídminnosti нуклеинови киселини аз отнех името им.
Структура на рибонуклеинова киселина
Първична структура на РНК
РНК молекулае линейни (т.е. недешифрирани) полинуклеотиди с подобен ДНК принцип на организация. РНК мономерите са нуклеотиди, които са съставени от фосфорна киселина, въглехидрат (рибоза) и азотна основа, свързани с 3", 5"-фосфодиестерни връзки. Полинуклеотидните копия на молекулата на РНК са полярни, т.е. 5'- и 3"-точките могат да бъдат нарушени. При това, на повърхността на ДНК, РНК е еднолентова молекула. Причината за тази иманентност са три характеристики на първичната структура:- РНК на повърхността на ДНК замества дезоксирибозата рибоза, тъй като може да има допълнителна хидроксидна група. Хидрокси група за разрушаване на двойната структура на по-малкия компакт
- В средата на някои глави или главни азотни основи (A, G, C и U) тиминът се заменя с урацл, който се отстранява от тимина без наличието на метилова група на 5-та позиция. В резултат на това силата на хидрофобното взаимодействие в комплементарната двойка A-U се променя, което също намалява ефективността на приемане на стабилни двойни молекули.
- Nareshti, РНК (особено tRNA) има висока zm_st t. второстепенни бази и нуклеозиди. Сред тях са дихидроуридин (в урацил, няма една подвариантна връзка), псевдоуридин (урацил, иначе, по-нисък, се свързва с рибоза), диметиладенин и диметилгуанин (в азотни субстанции, две допълнителни метилови групи) и много други. Mayzhe всички tsі podstavi scho nepromozhnі да поемат съдбата на допълващите vzaєmodіyah. По този начин метиловите групи в диметиладенина (на базата на тимин и 5-метилцитозин) са разположени при такъв атом, който в A-U двойката прави водни връзки; otzhe, сега tsey zv'yazyok zakryuchissya невъзможно. Продължаване на трансформацията на двойните молекули.
По този начин широко признатият склад на РНК и структурата на ДНК могат да имат голямо биологично значение: въпреки че функцията на една молекула на РНК може да се изпълнява само в рамка с един ланцет, което е най-очевидно за иРНК: важно е да се покаже, че молекула с двойно копие може да бъде полупрозрачна от рибозоми.
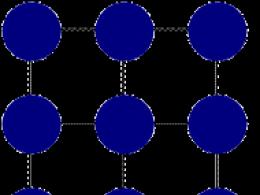
В същото време, затрупана от единична, в някои случаи ланцетната РНК може да образува бримки, издатини или „фиби“ с двойна структура (фиг. 1.). Tsya структура се стабилизира чрез размяната на бази в двойки A :: U и G ::: C. Въпреки това, могат да се правят "грешни" залози (например G U), а на определени места "фиби" и в огъня не се появяват по един и същи начин. В склада на такива бримки могат да бъдат редуцирани до 50% от общите нуклеотиди (особено в tRNA и rRNA). Общият брой на нуклеотидите в РНК варира от 75 единици до повече от хиляда. Алтернативно, намерете най-голямата РНК на порядък по-къса на хромозомна ДНК.
Първичната структура на иРНК е копирана от ДНК клетки, за да се извлече информация за първичната структура на полипептидния ланцет. Първичната структура на други видове РНК (тРНК, рРНК, рядка РНК) е остатъчно копие на генетичната програма на най-често срещаните ДНК гени.
Вторична и третична структура на РНК
Рибонуклеиновите киселини (РНК) са едноверижни молекули, следователно според вида на ДНК техните вторични и третични структури са неправилни. Qi структурите, които изглеждат като просторна конформация на полинуклеотиден ланцет, се образуват главно поради водно-свързани и хидрофобни взаимодействия между азотни бази. Докато спиралата е характерна за нативна ДНК молекула, структурата на РНК е разнообразна и лабилна. Рентгеновият дифракционен анализ показа, че черупките на полинуклеотидния РНК ланцет, огъвайки се, се навиват върху себе си с установените вътрешни спирални структури. Стабилизирането на структурите достига отвъд рахуните на комплементарни сдвоявания на азотните бази на антипаралелните ланци на ланцетника; специфични двойки тук е А-U, G-С і, повече, G-U. Гребените на тази молекула РНК се образуват като къси и дълги биспирални участъци, които лежат на едно копие; qі d_lyanki звучат като фиби. Моделът на вторичната структура на РНК с елементи, подобни на фиби, е създаден в края на 50-те години - в началото на 60-те години. ХХ чл. в лабораториите на А. С. Спирин (Русия) и П. Доти (САЩ).
| Deyaki vidi РНК | ||
| Види РНК | Rosemir в нуклеотиди | функция |
| gRNA - геномна РНК | 10000-100000 | |
| иРНК – информационна (матрична) РНК | 100-100000 | предава информация за структурата на протеина от молекулата на ДНК |
| тРНК - транспортна РНК | 70-90 | транспортира аминокиселини до мястото на протеиновия синтез |
| рРНК - рибозомна РНК | килка от дискретни класове от 100 до 500 000 | да се проведе в рибозомите, участвайки в подразделянето на рибозомната структура |
| sn-РНК - малка ядрена РНК | 100 | премахва интронита и ензимно свързва екзонита от иРНК. |
| sno-RNA - малка ядрена РНК | участие в директна или извършена модификация на бази в рРНК и малка ядрена РНК, като например метилиране и псевдоуридинизация. Повечето малки ядрени РНК се заместват от интрони на други гени | |
| srp-РНК - сигнал, който разпознава РНК | разпознава сигналната последователност на протеините, които се използват за експресия, и участва в пренасянето им през цитоплазмената мембрана | |
| mi-RNA - микро-РНК | 22 | контролират транслацията на структурни гени чрез комплементарно свързване към 3'-терминали на нетранслирани иРНК клетки |
Създаването на спирални структури е придружено от хипохромен ефект - промяна в оптичната плътност на РНК частиците при 260 nm. Ruinuvannya tsikh структури vídbuvaєtsya с намалена йонна сила на РНК разликата или дори нагряване до 60-70 ° C; Нарича се още топене и се обяснява със структурната преходна спирала - хаотична топка, която е придружена от увеличаване на оптичната дебелина на нуклеиновата киселина.
В клитините има видове РНК:
- информационна (или шаблонна) РНК (iRNA или mRNA) и нейната напред - хетерогенна ядрена РНК (rn-RNA)
- трансферна РНК (t-RNA) и нейната предна
- рибозома (r-RNA) и нейната предна част
- малка ядрена РНК (sn-РНК)
- малка ядрена РНК (сно-РНК)
- РНК за разпознаване на сигнала (srp-RNA)
- миРНК (ми-РНК)
- Митохондриална РНК (t + РНК).

Хетерогенна ядрена и информационна (матрична) РНК
Хетерогенната ядрена РНК е изключителна за еукариотите. Това е предната част на информационната РНК (i-RNA) за пренасяне на генетична информация под формата на ядрена ДНК към цитоплазмата. Хетерогенната ядрена РНК (премРНК) е оценена от радианския биохимик Г. П. Георгиев. Броят на видовете rn-РНК е равен на броя на гените, така че да служи като директно копие на последователностите, които кодират генома, след което може да има копия на палиндроми на ДНК, което е вторичната структура за отмъщение фиби и линейни дилянки. В процеса на транскрипция на РНК от ДНК ключова роля играе ензимът РНК полимераза II.
Информационната РНК се утаява в резултат на обработката (узряването) на rn-РНК, в която се раждат фиби, виждат се дивергенции (интрони), които не кодират, и кодиращите екзони са залепени заедно.
Информационната РНК (i-RNA) е копие на ДНК ДНК и изпълнява ролята на носител на генетична информация в ДНК преди синтеза на протеин (рибозома) и участва директно в нагъването на неговите молекули.
Зрялата матрична РНК има малък брой региони с различни функционални роли (фиг.)
- на 5"-края има така наречената "шапка" или капачка - дилянка от един-хотириокс модифицирани нуклеотиди. Такава структура защитава 5"-края на иРНК от ендонуклеази
- зад "cowpack" ide 5"-нетранслиран регион - последователност от десет десетки нуклеотиди. Той е комплементарен на една от другите три r-РНК, така че влиза в малката субединица на рибозомата. но не t излъчва себе си
- начален кодон – AUG, който кодира метионина. Всички иРНК имат един и същ начален кодон. Транслацията (четенето) на иРНК започва отново. Тъй като метионинът не е необходим за синтеза на пептидното копие, vin, като правило, се използва като част от N-края.
- Зад иницииращия кодон е следващата кодираща част, която трябва да извлече информация за последователността на аминокиселините в протеина. При еукариотите зрелите тРНК са моноцистронни, т.е. кожата от тях носеше информация за структурата само на един полипептиден ланцет.
Вдясно някои пептидни копия необичайно се установяват върху рибозоми и се разделят на цаца на други копия. Така че се използва, например, в синтеза на инсулин и цяла серия от олигопептидни хормони.
Кодиращата част на зрялата иРНК на еукариотите е пощадена от интрони - независимо дали има намесени последователности, които не са кодирани. С други думи, няма прекъсване в последователността на значещите кодони, тъй като трябва да се чете директно 5 " -> 3".
- След завършване на секвенционната последователност, кодонът на термина е известен - един от трите "мълчаливи" кодона: UAA, UAG или UGA (див. таблица на генетичния код по-долу).
- Cym кодонът може да бъде последван от 3'-нетранслиран регион, който значително измества 5'-нетранслирания регион надолу по веригата.
- Нарешти, може всички еукариотни иРНК (хистонови иРНК) на 3' края на поли(А)-фрагмента от 150-200 аденилови нуклеотида.
3'-нетранслационният ген и поли(А)-фрагментът могат да доведат до регулиране на продължителността на живота на иРНК, фрагментите на разрушаване на иРНК се контролират от 3'-екзонуклеази. След завършване на транслацията на иРНК, 10-15 нуклеотида се разделят на поли(А) фрагмента. Когато даден фрагмент е остарял, значителна част от иРНК започва да се свива (например 3'-нетранслирана клетка).
Общият брой на нуклеотидите в тРНК варира в брой хиляди. Когато tsimu от частта, която кодира, понякога могат да паднат по-малко от 60-70% от нуклеотидите.
В клитините молекулите на иРНК могат да бъдат свързани с протеини. Останете, очевидно, стабилизирайте линейната структура на иРНК, така че да изпревари светлината в частта, която кодира "фибичката". В допълнение, протеините могат да откраднат иРНК от предварително съсирване. Такива комплекси от иРНК с протеини понякога се наричат информозоми.
Транспортната РНК в цитоплазмата на клетката пренася аминокиселини в активирана форма към рибозомите, миризмите са прикрепени към пептидните копия в последователността на песента, която се задава от матрицата на РНК (mRNA). Към този момент има данни за нуклеотидната последователност на повече от 1700 вида tRNA от прокариотни и еукариотни организми. Всички миризми могат да бъдат комбинирани в първичната им структура, както и в метода на сгъване на полинуклеотидния ланцет във вторичната структура за комплементарните нуклеотидни обмени, които влизат преди тяхната структура.

Транспортната РНК в своя склад може да съдържа не повече от 100 нуклеотида, сред които има голям брой второстепенни или модифицирани нуклеотиди. Първата част от дешифрираната транспортна РНК беше РНК на була аланин, наблюдавана от дрожди. Анализът показа, че аланиновата РНК е съставена от 77 нуклеотида, смесени в строго определени последователности; техният склад включва така наречените второстепенни нуклеотиди, представени от атипични нуклеозиди
Alaninova tRNA отмъщава на 9 невъобразими бази с една или и двете декилком метилови групи, които се присъединяват към тях по ензимен път след установяването на фосфодиестерни връзки между нуклеотидите. Tsí podstavi nezdatnі до приемането на традиционните двойки; Възможно е да служи за това, за това, pereshkati sparjuts на основите на молекулите на молекулите на молекулата на молекулата на имението на посочените мили, яки е вторичната навигация на РНК, на борда, ензимите, нестареещите . Последователността на нуклеотидите в тРНК по същество означава, че същата последователност се наблюдава в гените, в които се синтезира тРНК. Последователността може да бъде въведена въз основа на правилата за специфично сдвояване на подстанции, установени от Watson и Crick. През 1970 г. rocibula синтезира една единствена двойно живееща ДНК молекула с двойна последователност от 77 нуклеотида и изглежда, че тя може да бъде матрица за индуциране на аланин трансферна РНК. Точно така, първата част от синтезирането на ген. |

Транскрипция на тРНК
Транскрипцията на t-RNA молекули е подобна на последователностите, които я кодират в ДНК с участието на ензима РНК полимераза III. В резултат на транскрипцията първичната структура на тРНК се формира като линейна молекула. Образуването започва от нагъването на последователността от нуклеотиди от РНК полимераза по подобен на гена начин, за да се премахне информацията за дадената трансферна РНК. Тази последователност е линейна полинуклеотидна последователност, в която нуклеотидите вървят един по един. Линейното полинуклеотидно копие е първичната РНК, предната част на тРНК, която включва интрони - неинформативни излишни нуклеотиди. В тази група организацията на пре-tRNA не е функционална. Utvoryuyuchisya в различни части на ДНК на хромозомите pre-tRNA, за да замени излишъка от приблизително 40 нуклеотида по двойки със зряла tRNA.
Друг етап от новосъздадения фронт на тРНК е да претърпи посттранскрипционно съзряване или обработка. В хода на обработката неинформативните излишни пре-РНК се отстраняват и се създават възрастни функционални РНК молекули.
пре-тРНК обработка
Обработката се инициира от факта, че вътрешномолекулните водни връзки са установени в транскрипта и молекулата на тРНК е оформена като стабилно листо. Това е втората линия на тРНК организация, в която тРНК молекулата все още е функционална. По-нататъшно развитие на неинформативни клетки от пре-РНК, растеж на информативни клетки на "счупени гени" - сплайсинг и модификация на 5 "- и 3"-вид клетки на РНК.
Развитието на неинформативни пре-РНК клетки се поддържа от рибонуклеази (екзотични ендонуклеази). След отстраняването на излишните нуклеотиди настъпва метилиране на тРНК бази. Реакцията се медиира от метилтрансферази. S-аденозилметионинът действа като донор на метилови групи. Обръщане на метилирането на разрушаването на тРНК от нуклеази. Остатъчно зрялата tRNA се утаява по път за пристигането на специфична тройка нуклеотиди (акцепторен край) - CCA, която се създава от специална РНК полимераза.
След завършване на обработката във вторичната структура, допълнителни водни връзки за ребрата на някои тРНК отново се установяват, за да преминат към третото ниво на организация и да изглеждат като така наречената L-форма. Този вид тРНК изглежда като хиалоплазма.
Будова тРНК
Структурата на транспортната РНК се основава на верига от нуклеотиди. Въпреки това, чрез тези, които, било то копие от нуклеотиди, могат да бъдат положително и отрицателно заредени части, вината не могат да бъдат в клетка в бучещо състояние. Заредените части, привлечени една към една, лесно установяват водни връзки помежду си, следвайки принципа на взаимното допълване. Водните връзки химерно усукват t-RNA веригата и я навиват в това положение. В резултат на това вторичната структура на t-РНК може да изглежда като "стабилен лист" (фиг.), Който отмъщава за 4 двуклюни лози в структурата си. Голям брой второстепенни и модифицирани нуклеотиди, които се намират в ланцетите на тРНК и късно до комплементарни взаимодействия, образуват 5 еднопосочни котила.
вкл. Вторичната структура на t-RNA се установява след интра-лантерно сдвояване на комплементарни нуклеотиди в четири tRNA клетки. ТРНК клетките, които не излъчват до установяването на водни връзки между нуклеотидите, установяват бримки или линейни връзки. В тРНК се наблюдават следните структурни разделения:
- Акцептор дилянка (кинец)която е съставена от няколко линейни разширения на нуклеотиди, три от които могат да имат една и съща последователност във всички видове tRNA - CCA. Добавя се хидроксил 3 "-ВІН към аденозин. Карбоксилната група на аминокиселината се добавя към новата и името на tRNA семейството е акцепторий. Свързвайки се с 3"-хидроксилната група на аденозин, аминокиселината tRNA е доставени за синтеза на рибозоми.
- Антикодонна примказвук utvoryuєtsya simoma нуклеотиди. Има триплет от нуклеотиди, специфични за tRNA на кожата, който се нарича антикодон. Антикодонът на tRNA, следвайки принципа на комплементарност, се свързва с кодона на mRNA. Взаимодействието кодон-антикодон определя реда на разпределяне на аминокиселините в полипептидния ланцет по време на нагъване в рибозомите.
- Псевдоуридилна примка (или TΨC-примка), който се състои от седем нуклеотида и obov'yazykovo отмъщение излишък псевдоуридил киселина Допуска се, че псевдоуридил цикълът участва в свързването на тРНК от рибозомата.
- Дихидроурид или D-контур, който се състои от 8-12 нуклеотидни излишъци, сред които има някои излишъци на дихидроуридин. Важно е да се отбележи, че D-контурът е необходим за свързване с аминоацил-тРНК синтетазата, тъй като участва в разпознатата аминокиселина на собствената си тРНК (разделение "Биосинтеза на протеин"),
- Додатков цикъл, Yaka varіyyuê за razmirami и запасни нуклеотиди в различни tRNA.
Третиновата структура на t-RNA вече няма формата на стабилен лист. За установяване на водни връзки между нуклеотиди от различни части на "аркушите конюшни" пелусите се извиват върху тялото на молекулата и се установяват в тази позиция на допълнителните връзки на Ван дер Ваалс, което предполага формата на буквата G или L , Наличието на стабилна третична структура е още една характеристика на t-RNA, показателна за два линейни m-RNA полинуклеотида. За да се разбере как различните части на вторичната структура на t-RNA с установена третична структура могат да се видят на фиг., като се покаже цветовата схема на вторичната структура на третичната структура на t-RNA.


Транспортна РНК (t-RNA) за пренасяне на аминокиселини от цитоплазмата до рибозомата по време на протеиновия синтез. От таблицата с генетичния код се вижда, че кожната аминокиселина е кодирана от десетични последователности от нуклеотиди, така че кожните аминокиселини дават своята транспортна РНК. В резултат на това разнообразието на t-RNA е голямо: от един до шест вида за кожа с 20 аминокиселини. Вижте тРНК, които се свързват с една и съща аминокиселина, се наричат изоакцептори (например аланинът може да бъде прикрепен към тРНК, антикодон, който ще допълни кодоните GCU, GCC, GCA, GCG). Специфичността на tRNA се обозначава с горния индекс, например: tRNA Ala.
За процеса на протеинов синтез от главните функционални части на t-RNA е: антикодон - последователност от нуклеотиди, които са разположени на антикодоновата верига, комплементарна на кодона на информационната РНК (i-RNA) и акцепторната част - дължина на антикодона на края на t-РНК, към който е добавена аминокиселина. Последователността на базите в антикодона може директно да се депозира под формата на аминокиселина, прикрепена към 3"-кинца. в основата на предаването на генетична информация, носеща такава t-RNA.
В процеса на синтез на т-РНК протеина, антикодонът разпознава трибуквената последователност на генетичния код (кодона) на i-РНК, който представлява една аминокиселина, прикрепена към другия край на тРНК. Само в случай на различна комплементарност на антикодона към иРНК плочата на трансферната РНК той може да бъде добавен към нея и да даде аминокиселината, която да бъде пренесена в матрицата на протеиновото копие. Взаимодействието на t-RNA и i-RNA се открива в рибозомата, тъй като тя също е активен участник в транслацията.
Разпознаването на t-RNA на нейната аминокиселина и кодон на t-RNA се определя от реда на песните:
- Свързването на "собствени" аминокиселини с t-RNA зависи от допълнителен ензим - специфична аминоацил-tRNA синтетаза
Голямото разнообразие от аминоацил-тРНК-синтетази се дължи на броя на тРНК, които са заместени с аминокиселини. Те се наричат накратко ARSazi. Аминоацил-тРНК синтетазите са големи молекули (молекулно тегло 100 000 - 240 000), които образуват четвърт структура. Те знаят точно за tRNA и аминокиселините и ги катализират една до друга. За TSO, ATF е процесът на ATF, vicoristovye на Активиране на карбоксид Kinzili I Podnnannya към Gidroxilu (3 "-on) аденозин на акцептора Kintsya (TsCA) TRNA три центъра на ATF: свързване с установяване на ковалентна връзка, tRNA-подобни аминокиселини и хидролиза на такава връзка в различни несъответствия (прикрепване към tRNA "не tієї" аминокиселини).
ARSase може да бъде в състояние да вибрира победоносно с разпознаването на диапазона от tRNAs за кожни аминокиселини, tobto. аминокиселината се въвежда от жична ланка и нейната собствена tRNA се извежда пред нея. Нека тРНК чрез проста дифузия пренася аминокиселината, донесена до нея, до рибозомите, разлагайки нагъването на протеина от аминокиселини, които трябва да се намират в различни аминоацил-тРНК.

Свързване на аминокиселини с тРНК
Свързването на tRNA и аминокиселините следва този ред (фиг.): аминокиселина и ATP молекула се добавят към аминоацил-tRNA синтетазата. За леко амино насочване, ATP молекулата вибрира енергия, разделяйки две фосфатни групи. Останалият АМФ (аденозин монофосфат) се присъединява към аминокиселините, подготвяйки се за съединяване с акцепторната пластина на тРНК - акцепторната фибичка. След тази синтетаза същата аминокиселина на тРНК на спората идва на себе си. На този етап се извършва повторна проверка на вида на тРНК синтетазата. В различните видове tRNA тя е силно прикрепена към синтетазата, променяйки структурата си, което води до стартиране на процеса на амино-насочване - добавяне на аминокиселини към tRNA.
Аминоацилирането се дължи на процеса на заместване на AMP молекулата, добавена към аминокиселината с tRNA молекулата. След замяната на AMP, синтетазата се изчерпва и tRNA се подрязва за останалата част от преобразуването на аминокиселината.
Повторна проверка на типа tRNA на добавената аминокиселина
Синтетазният модел за обръщане на tRNA типа на добавените аминокиселини предава наличието на два активни центъра: синтетичен и коригиращ. В синтетичния център tRNA се добавя към аминокиселината. Акцепторната клетка на тРНК, уловена от синтетаза, е в контакт със синтетичния център на кочана, където се намира аминокиселината и се получава с AMP. Този контакт на акцепторната клетка на tRNA поражда неестествен вигин преди добавянето на аминокиселини. Освен това, тъй като аминокиселината се добавя към акцепторната клетка на tRNA, е необходимо тази клетка да се намира в синтетичния център през есента, tRNA се изправя и се премества към нея от аминокиселината до коригиращия център. Когато молекулата на аминокиселината се смеси, тя се добавя към тРНК, но се добавя към корекционния център, аминокиселината се разпознава като неправилна и влиза в тРНК. Синтетазата е готова за следващия цикъл. Когато молекулата на аминокиселината е разширена, прикрепена към тРНК, тогава коригиращият център се зарежда с аминокиселината, тРНК се активира: тя е готова да играе роля в транслацията на протеина. И синтетазата е готова да приеме нови аминокиселини и тРНК и да започне втори цикъл.
Средният брой несинтетични аминокиселини със синтетазата се среща в 1-вата капка от 50 хиляди, а с tRNA на tRNA само веднъж на 100 хиляди дни.
- Взаимодействието между тРНК кодон и тРНК антикодон следва принципа на комплементарност и антипаралелизъм
Взаимодействието на тРНК с иРНК кодона съгласно принципа на комплементарност и антипаралелизъм означава: сенс кодонът иРНК чете на права линия 5 "-> 3", тогава антикодонът в тРНК е виновен за четене на права линия 3 "-> 5" . При тези първи две сдвоявания кодонът и антикодонът са строго допълващи се, така че сдвояването на A U и G C не е възможно. Допустимите залози се определят от схемата:
Излизат три схеми.
- Молекулата на tRNA се свързва само с първия тип кодон, като третия нуклеотид в нейния антикодон - C или A
- tRNA се появява с два вида кодони, така че антикодонът завършва на Y или R.
- I, nareshti, tRNA се появява с три вида кодони, така че антикодонът завършва с I (инозинов нуклеотид); такава ситуация, zocrema, в аланинова тРНК.
Zvídsi на свой ред крещи, че за разпознаването на 61 сетивни кодона по принцип не е същото, а по-малък брой различни тРНК.
Рибозомна РНК

Рибозомната РНК е в основата на образуването на субединици на рибозоми. Рибозомите осигуряват пространство за взаимно замразяване на тРНК и тРНК в процеса на синтез на протеини.
Кожната рибозома е изградена от големи и малки субединици. Субединиците включват голям брой протеини и рибозомна РНК, които не изискват транслация. Рибозомите, както и рибозомната РНК, се отличават с коефициента на утаяване (седиментация), който се измерва в единици Svedberg (S). Tsey коефициент за отлагане според стабилността на утаяване на субединици по време на центрофугиране в богата водна среда.
Кожната рибозома на еукариотите има коефициент на утаяване, равен на 80S, и е прието да се обозначава 80S-частица. Вон включително
- малка субединица (40S) за компенсиране на рибозомната РНК с коефициента на утаяване на 18S рРНК и 30 молекули от различни протеини,
- голяма субединица (60S), която включва 3 различни рРНК молекули (една дълга и две къси - 5S, 5.8S и 28S), както и 45 протеинови молекули.
Субединицата utvoryuyut "скелет" на рибозомата, кожата на някои от ексудатите с техните протеини. Коефициентът на утаяване на общата рибозома не се променя от сумата на коефициентите на две субединици, което е свързано с пространствената конфигурация на молекулата.
Прикрепването на рибозомите на прокариотите и еукариотите е приблизително еднакво. Смърдят с по-малко молекулно тегло. Бактериалната рибозома има коефициент на утаяване 70S и е обозначена като 70S-част, което показва по-малко утаяване; отмъсти си
- малка (30S) субединица - 16S рРНК + протеини
- голяма субединица (50S) - 23S rRNA + 5S rRNA + протеини на голямата субединица (малка)
В рРНК на средните азотни бази звучи по-ниско вместо гуанин и цитозин. Малките нуклеозиди също се улавят, но не толкова често, колкото в тРНК: приблизително 1%. Ce, главно нуклеозиди, метилиране на рибоза. Във вторичната структура на рРНК има много клетки-джуджета и бримки (малки). Това е вид РНК молекули, които се установяват в два последователни процеса - ДНК транскрипция и узряване (обработка) на РНК.
rRNA транскрипция от ДНК и обработка на rRNA
PrerRNA се установява в ядрото, където се намира транскрипцията на rRNA. Транскрипцията на рРНК от ДНК се поддържа от две комплементарни РНК полимерази. РНК полимераза I транскрибира 5S, 5.8S и 28S в един дълъг 45S транскрипт, който след това се подразделя на необходимите части. По този начин се осигурява равен брой молекули. При хората кожният хаплоиден геном съдържа приблизително 250 копия на ДНК последователността, кодираща 45S транскрипта. Вонята е подредена в пет клъстерирани тандемни повторения (т.е. един по един по двойки) в късите рамена на хромозоми 13, 14, 15, 21 и 22. Тези клетки са ядрени организатори, както и тяхната транскрипция и по-нататъшна обработка на 45S-транскрипта на ядрото.
Не по-малко от три групи от хромозома 1 с 2000 копия на 5S-pRNA гена. Їx транскрипцията протича в присъствието на РНК полимераза III ниво на ядрото.
Повече от половината от пре-рРНК се губи в процеса на обработка и рРНК узрява. Някои от нуклеотидите на рРНК подлежат на модификация, която е подобна на метилираните бази. Реакцията се медиира от метилтрансферази. S-аденозилметионинът играе ролята на донор на метални групи. РРНК са свързани в ядрото с рибозомни протеини, които се намират тук в цитоплазмата и образуват малки и големи подчасти на рибозомата. Видовете rRNA се транспортират от ядрото до цитоплазмата в комплекс с протеин, което допълнително ги предпазва от разрушаване и пренасяне.
рибозомен центр
Рибозомите се намират естествено в други органели на клетката. В цитоплазмата миризмите се чуват в две състояния: в непрактичното, ако субединицата на водата е голяма и малка, едно в едно, а в активната функцията на протеиновия синтез е нарушена, ако субединиците са един с един.
Процесът на сглобяване на субединици на рибозоми или сгъване на активни рибозоми е посочен като начало на транслацията. Тази селекция е строго подредена, за да се осигурят богатите функционални центрове на рибозомите. Всички центрове са разположени на повърхностите, където контактуват, двете субединици на рибозомата. Пред тях се виждат:
- иРНК свързващ център (М-център). В смилането на 18S рРНК, което се допълва от 5-9 нуклеотидно разтягане в 5'-нетранслирания иРНК фрагмент
- Пептидил център (Р-център). В началото на процеса на транслация той е свързан с инициираща aa-tRNA. При еукариотите първоначалният кодон на всички иРНК винаги кодира метионин, който е една от двете метионинови аа-тРНК, което се обозначава с долния индекс i: Met-tRNA i Met. В напредналите етапи на транслацията пептидил-тРНК се намира в Р-центъра, който вече е синтезирана част от пептида lanciug.
Може да се говори и за Е-център (като "изход" - изход), където се движи тРНК, която е включила връзка с пептидила, преди да напусне рибозомата. Но центърът може да се разглежда като складов P-център.
- Аминокиселинният център (А-център) е мястото на свързване на черната aa-tRNA.
- Пептидилтрансферазен център (PTF център) - катализира преноса на пептидил от склада на пептидил-тРНК към черната аа-тРНК, която се намира в А центъра. В този случай се образува още една пептидна връзка и пептидите се включват от една аминокиселина.
Както в аминокиселинния център, така и в пептидиловия център, антикодонната верига на видпоидната тРНК (аа-тРНК или пептидил-тРНК), очевидно, е удължена до М-центъра - свързващият център на матричната РНК (връзка с иРНК) и е акцептор към PTP центъра.
Rozpodíl центрове между субединици
Разделението на центровете между субединиците на рибозомата е както следва:
- Малка субединица.Ако не е възможно 18S-rRNA да си отмъсти, тъй като иРНК се свързва с мястото, тогава центърът се намира в тази субединица. Освен това има основната част от А-центъра и малка част от Р-центъра.
- Страхотна подединица. На нейната контактна повърхност на фрезоването на другите части на P-i A-центровете. В случая на Р-центъра има цялата основна част, а в случая на А-центъра има връзка на акцепторната верига AA-tRNA с аминокиселинния радикал (аминоацил); reshta w и повече aa-tRNA се появяват с минимална субединица. Голямото подразделение също се намира в центъра на PTF.
Иницииране на рибозома (подготовка на рибозома преди протеинов синтез)
Синтезът на протеина, или в хода на транслацията, е разделен на три фази: започване (кочан), удължаване (понижаване на полипептидното копие) и терминиране (завършване). Във фазата на започване започва подготовката на рибозомата за работа: следващата субединица. В бактериалните и еукариотните рибозоми субединицата и транслационното ухо протичат по различен начин.
Началото на превода е най-важният процес. В новия крем рибозомните субединици, тРНК и тРНК поемат ролята на GTP и три протеинови фактора на иницииране (IF-1, IF-2 и IF-3), които не са складови компоненти на рибозомата. Иницииращите фактори улесняват свързването на иРНК към малката субединица и GTP. GTP за хидролиза на rahunok осигурява енергия за процеса на zmikannya субединици на рибозомата.

- Инициирането се дължи на факта, че малка субединица (40S) изглежда се дължи на иницииращия фактор IF-3, което води до изместване към предната връзка на голямата субединица и възможността за прилепване към нея иРНК.
- По-далеч от комплекса "малка субединица (40S) + IF-3", е прикрепена иРНК (неговият 5"-нетранслиран спейсер).
- В допълнение към комплекса "малка субединица + IF-3 + mRNA" се добавят още два иницииращи фактора: IF-1 и IF-2, като останалите носят специална трансферна РНК, която се нарича инициираща aa-tRNA. Комплексът включва и ГТФ към склад.
Малка субединица след иРНК представлява два кодона за четене. Към първия IF-2 протеин е прикрепена инициаторната aa-tRNA. Друг кодон затваря протеина IF-1, който го блокира и не му позволява да достигне до атакуващата тРНК, докато рибозомата не бъде пренагъната.
- Тъй като свързването на иницииращата aa-tRNA, след това Met-tRNA i Met за комплементарното взаимодействие с иРНК (първоначален кодон AUG) се вмъква на собственото си място в P-центъра, свързването на субединиците на рибозомата е установени. GTP се хидролизира до GDP и неорганичен фосфат и енергията, която се наблюдава по време на развитието на тази макроергична връзка, създава термодинамичен стимул за процеса да протече по необходимия директен начин. В същото време иницииращите фактори лишават рибозомата.
По този начин се оформя своеобразен „сандвич“ от някои от основните компоненти. Когато това се случи, иницииращият кодон на иРНК (AUG) се появява в Р-центъра на рибозомната рибозома и към него се свързва аа-тРНК, която инициира. Ролята на пептидил-тРНК играе ролята на пептидил-тРНК, когато се установи първата пептидна връзка.

РНК транскриптите, синтезирани с помощта на РНК полимераза, започват да разпознават далечни ензимни трансформации, наречени посттранскрипционна обработка, и само малко от смрадта придобиват своята функционална активност. Транскрипти на незряла информационна РНК се наричат хетерогенна ядрена РНК (hnRNA). Вонята е съставена от сбор от още две РНК молекули, които отмъщават на интрона и екзониума. Съзряването (обработката) на hnRNA в еукариотите включва набор от етапи, един от които включва разделяне на интрони - нетранслирани вмъкнати последователности и сливане на екзони. Процесът протича по такъв начин, че екзониумът един по един се поставя, така че кодиращите иРНК фрагменти не са физически разделени. Exoni определено са свързани помежду си за помощ на молекули, тъй като те се наричат малки ядрени РНК (snRNA). Функцията на тези къси ядрени РНК, които са съставени от приблизително 100 нуклеотида, отдавна е неясна. Тя е далеч, за да установи след това, както беше показано, че тяхната нуклеотидна последователност е комплементарна на последователностите в краищата на z интроните на кожата. В резултат на сдвояването на базите, които се срещат в snRNA и в краищата на интрона с примка, последователностите на двата екзона се приближават по такъв начин, че става възможно разделянето на интрона, разделянето им и ензимното снаждане (кодиране фрагменти). По този начин snRNA молекулите играят ролята на тимхазиеви матрици, които са близо до един в единия край на два екзона, за да може сплайсингът да се случи в правилното място (фиг.).
Трансформацията на hnRNA в iRNA преминава през пътя на отдалечените интрони в ядрения комплекс от РНК-протеини, наречен сплайсома. Кожната сплайсома има ядро, което се състои от три малки (с ниско молекулно тегло) ядрени рибонуклеопротеини или snurpiv. Skin snurp, за да отмъсти за една малка ядрена РНК и цаца бели. Има стотици различни малки ядрени РНК, които се транскрибират в основната РНК полимераза II. Важно е да се отбележи, че основната им функция е разпознаването на специфични рибонуклеинови последователности за допълнително сдвояване на бази за типа РНК-РНК. За обработката на hnRNA най-важните са Ul, U2, U4/U6 и U5.
Митохондриална РНК
Митохондриалната ДНК е непрекъсната верига и кодира 13 полипептида, 22 tRNA и 2 rRNA (16S и 23S). Повечето от гените са разположени на едно (важно) копие, протеята на копието е скрита и на допълващия бял дроб. В този случай ланцетните се транскрибират в привидно непрекъснати транскрипти за помощта на специфична за митохондриите РНК полимераза. Този ензим е кодиран от ядрения ген. След това всички молекули на РНК се разделят на 37 вида и тРНК, рРНК и тРНК транслират съвместно 13 тРНК. Има голям брой адитивни протеини, които трябва да достигнат до митохондриите от цитоплазмата и се транслират от ядрените гени. При пациенти със системни червеи се откриват антитела срещу snurp-протеините на тялото. Освен това е важно да се отбележи, че големият брой гени в малката ядрена РНК на хромозома 15q играе важна роля в патогенезата на синдрома на Прадер-Уили (намаляване на олигофренията, нисък растеж, затлъстяване, хипотония на m'yaziv ).
Има три основни функционални типа РНК в цитоплазмата на клитина:
- матрична РНК (иРНК), която определя функциите на матрици за синтез на протеини;
- рибозомна РНК (рРНК), която представлява ролята на структурните компоненти на рибозомите;
- транспортна РНК (тРНК), която транслира (транслира) иРНК информацията в последователността от аминокиселини на белтъчната молекула.
В ядрото на клитините се открива ядрена РНК, която варира от 4 до 10% от общата клетъчна РНК. Основната маса на ядрената РНК е представена от високомолекулни прекурсори на рибозомни и трансферни РНК. Високомолекулните прекурсори на рРНК (28 S, 18 S и 5 S РНК) са локализирани предимно в ядрото.
РНК д основен генетичен материалв някои вируси, животни и розлин (геномна РНК). Повечето РНК вируси се характеризират с обратна транскрипция на техния РНК геном, която се насочва от обратна транскриптаза.
Всички рибонуклеинови киселини рибонуклеотидни полимери,свързани, както в молекулата на ДНК, чрез 3,5"-фосфородиестерни връзки. От вида на ДНК, която има двойна структура, РНК еднолентови линейни полимерни молекули.
Budov иРНК. иРНК е най-хетерогенният клас РНК по отношение на стабилност и стабилност. Количеството иРНК в клитините става 2-6% от общата РНК. иРНК са изградени от клетки - цистрони, които определят последователността на аминокиселините в протеините, които са кодирани от тях.
Будова тРНК . Транспортните РНК определят функциите на медиаторите (адапторите) за часа на транслацията на иРНК. Приблизително 15% от общата клетъчна РНК попада в тази пропорция. Кожата от 20 протеиногенни аминокиселини има своя собствена тРНК. За активни аминокиселини, които са кодирани от два или повече кодона, се използва tRNA снаждане. tRNA са относително малки едноланцетни молекули, които се състоят от 70-93 нуклеотида. Молекулното тегло става (2.4-3.1) .104 kDa.
Вторична структура на тРНКобразувани за rahunok на максималния брой водни връзки между вътрешните молекулни комплементарни двойки азотни бази. В резултат на установяването на тези връзки, полинуклеотидният ланцет на тРНК се усуква около установените спирализации, които завършват в бримки от несдвоени нуклеотиди. Разширяване на изображения на вторичните структури на usíh tRNA формата. стабилен лист.
При "стабилния чаршаф" те се разделят chotiri obov'yazkovі hіlki, повече dovgі tRNA, освен това, отмъщение кратко p'yat (допълнително) gílku. Адаптерната функция на тРНК се осигурява от акцепторен ръкав, до 3 "-точка, който е свързан с етерна връзка с излишък от аминокиселини, а акцепторният ръкав се противопоставя на антикодонов ръкав, в горната част на който има примка, която отмъщава на антикодона.код
T-gylka, която носи псевдоуридинова верига (TyC-loop), осигурява взаимодействието на tRNA с рибозомите.
D-gylka, която носи дехидроуридиновата верига, осигурява взаимодействието на тРНК с жизнеспособната аминоацил-тРНК синтетаза.
Вторична структура на тРНК
Функциите на петия придатък гилка все още са малко проучени, а по-скоро разликата между различните tRNA молекули.
Третинова структура на тРНКтой е по-компактен и се урежда чрез пътя на сближаване на okremyhs на стабилния лист за лена на допълнителните водни връзки с разтворите на L-подобната структура "Liktovogo zginu". С кое акцепторно рамо, което свързва аминокиселината, разпространяваме в единия край на молекулата, а антикодона в другия.

Третинова структура на tRNA (от A.S. Spirin)
Budov rRNA и рибозома . Рибозомните РНК формират основата, с която специфични протеини се свързват един с друг по време на установяването на рибозомите. Рибозоми- Ce нуклеопротеинови органели, които осигуряват синтеза на протеин върху иРНК. Броят на рибозомите в klitz е още по-голям: от 104 при прокариотите до 106 при еукариотите. Те са локализирани от рибозоми в ранга на главата в цитоплазмата, при еукариотите, освен това в ядрото, в матрицата на митохондриите и в стромата на хлоропластите. Рибозомите са изградени от две подчасти: голяма и малка. По големина и молекулна маса всички рибозоми се разделят на 3 групи - 70S рибозоми на прокариотите (S-коефициент на седиментация), които са съставени от малки 30S и големи 50S субчасти; 80S рибозоми на еукариоти, които са съставени от 40S малки и 60S големи субединици.
Малък подразделРибозомата 80S е изпълнена с една молекула рРНК (18S) и 33 молекули от различни протеини. Страхотен подраздел trioma се състои от rRNA молекули (5S, 5.8S и 28S) и приблизително 50 протеина.
Вторична структура на рРНК utvoryutsya зад къси двойноверижни участъци на молекула - фиби (около 2/3 от rRNA), 1/3 - е представена от един-важни парцели, богат на пуринови нуклеотиди.
Помощник. Независимо от тези, че tRNA е значителна, rozpovid за нейната budova, особено за функционирането, заслужено на okremiya rasdіl.
По-късно tRNA е „адаптер“, който с единия си край разпознава трилитерната последователност на генетичния код, което го прави една единствена аминокиселина, прикрепена към другия край на tRNA. Има 3 нуклеотида в горната част на информационната РНК и в края на транспортната РНК антикодон. Само във времена на комплементарност на антикодона към иРНК носителят на трансферната РНК може да стигне до него. Алтернативно, тРНК не може да влезе в иРНК сама, тя се нуждае от помощта на рибозомата, тъй като тя е мястото на тяхното взаимодействие, а също и активен участник в транслацията. Например, самата рибозома създава връзки между аминокиселините, донесени от тРНК, образувайки протеиново копие.
Особеностите на тРНК са маркирани с генетичен код, така че протеинът зад гена да управлява, така че транспортната РНК да бъде разчетена. Този код работи при кожната болест, която живее на Земята: създаването на вируса е записано от същите трилитерни кодони, които са написани, за да записват „инструкциите за сгъване“ на делфина. Експериментално е проверено, че гените на една жива природа, поставени в клетките на друга, се копират и транслират в протеини, сякаш не приличат на гени, които се създават в клетките на владетеля. На единството на генетичния код се основава производството на колонии от модифицирани чревни пръчици от инсулин и богатство от други човешки ензими, които са победоносни като букви за хора, чийто организъм не е жизнеспособен или липсва. Независимо от очевидната разлика между човешката и чревната пръчка, човешките протеини лесно се създават зад йога столове за помощта на чревната пръчка. Не е изненадващо, че транспортните РНК от различни източници са още по-малко разпознати.

Кожен кодон от този списък стоп кодони, които дават сигнал за завършване на транслацията, могат да бъдат разпознати чрез трансферна РНК. Разпознаването е необходимо за допълнителна помощ при влизането на матрична РНК в антикодон, който може да се асоциира само с един кодон от списъка, че тРНК може да разпознае само един кодон. Освен това в клитината има само 61 вида от тези молекули. Всъщност си струва повече, защото в определени ситуации за четене на матрична РНК не е достатъчно само да се използва майчиният антикодон: необходимо е да се търсят други умове, вероятно е създадена специална модифицирана тРНК.
На пръв поглед риосноманността на TRNA майо е оценката на процеса на превод: Ajjnu Zikh молекули на превода на пиротите на рибозомния кодон на матричната РНК към неговия антикодон - чудото на механичния заек, мандатът на мандата на заека, мандата И в резултат на еволюцията се формираха същите клинични механизми, които биха помогнали на тези проблеми. Например, броят на тРНК на дермалните видове в клетките показва, че често аминокиселината, която не присъства във вида, е вградена в протеините, които ще бъдат. Є аминокиселини, които рядко са победни и често победни, а броят на неносещите тРНК е същият, което значително улеснява сгъването на протеините. Следователно има малко "релевантни" аминокиселини и подобни тРНК в клитините, а тези, които често се увеличават, вибрират в голям брой.

С такъв набор от tRNA молекули всички миризми са повече или по-малко сходни, което, като се погледнат ежедневните им функции, е най-малко по-малко тежко за всички видове ориз. Ако погледнете тривимичната схема на тРНК, ще видите огромна купчина атоми. Известно е, че молекулата на отримана се сгъва в резултат на дълго ланцетно нуклеотидно сгъване, но самата тя го прави.
Възможно е да се извърши етапът на нейното приемане, като се започне от първия: сгъването на последователността от нуклеотиди от РНК полимераза е подобно на гена, за да се премахне информацията за дадената трансферна РНК. Редът на последователност един по един от нуклеотидите и техният брой се нарича първична структура на тРНК. Изглежда, че първичната структура на тРНК е кодирана в гена, разчетен от РНК полимераза. Вместо това първичната структура е последователност от еднакво прости молекули от един и същи тип, включително нагъната, нагъната полимерна молекула. Например, първичната структура на протеинова молекула е проста последователност от складови аминокиселини.

Без значение колко ланцет от нуклеотиди, не можете да бъдете в клише при ревящ стан, просто като се разтегнете на линията. По краищата на нуклеотидите вече има богато положително и отрицателно заредени части, които просто образуват водни връзки една по една. За тези, как се установяват едни и същи връзки между нуклеотидите на две ДНК молекули, свързвайки ги със спиралата на основата, е описано в , а за подробности относно водните връзки можете да отидете на. Водните връзки са по-малки, по-ниски връзки между атомите в молекулите, но все пак е достатъчно да се усуче тРНК и да се подстриже химерно в тази позиция. Гръбнакът на връзките е уреден само с няколко активни нуклеотида, сгъвайки тРНК във фигурата върху щифта на стабилния лист. Резултатът от което се нарича кочан ларинкс вторична структура тРНК. От схемата на zliva става ясно, че активните нуклеотиди са свързани с водни връзки, в противен случай се запълват с несдвоени, образувайки пръстени и бримки. Разликите между вторичната структура на различните видове тРНК са свързани с разликите в тяхната първична структура. Това се проявява в различната дължина на "стабилните листа" или "стъблото" за перата на различната дължина на външния ланцет от нуклеотиди.
Втората разлика в първичната структура на различните тРНК е тези, които имат еднакви нуклеотиди в определени позиции на вонята (на диаграмата, в хоризонталните линии, те са обозначени с първите букви от техните имена), по-големият брой нуклеотиди в различни тРНК се пресичат. Въведена е по-обща схема за всички тРНК, така че обсъжданите нуклеотиди са с номера.
Главните функционални части на тРНК е:
=) антикодон, така че последователността от нуклеотиди е комплементарна на единичен кодон на матрична РНК, който се намира на антикодон фиби
=) акцепторни кинети, До каква степен може да се добави аминокиселината. Vіn znahoditsya protilezhny страна vіd antikodonovoї фиби.
Всъщност оригиналната tRNA не изглежда така, както изглежда на схемата на вторичната структура, така че за целите на нейното установяване са събрани само няколко нуклеотида, а reshta са оставени несдвоени. За уреждане на водни връзки между нуклеотиди от различни части на листа на конюшнята, вените се извиват нагоре в богато нагънати. третична структурапод формата на буквата L. Можете да разберете, че самите части на вторичната структура са били огънати за приемане на третичната, като поставите цветовете на техните диаграми по-долу. Антикодонната фибичка, маркирана с черни и бели цветове, е оставена в долната част (брадавица на паметта, това, което е „долу“, трябва да се разбере: визуално изобразете самата tRNA в такава просторна ориентация в схемите за транслация на протеини), и акцепторният връх (жълт пръстен) е огънат.

Самата tRNA изглежда така, сякаш е готова преди добавянето на аминокиселини. Не е възможно самоасоцииране с аминокиселина tRNA, за което е необходимо участието на специален ензим: аминоацил-тРНК синтетаза. Броят на видовете синтетаза в clitiny варира в зависимост от броя на видовете tRNA.
Единството на формите на всички видове тРНК е необходимо, за да може рибозомата да разпознае дали са, да приеме техните пръчки с иРНК, да се премести от едно място на друго. Различните видове тРНК се променят спорадично един по един, което прави работата на рибозомата по-сложна, критично променяйки скоростта на протеиновия синтез. По този начин естествените видове се насочват към тези, които правят тРНК подобни една на друга. Но в същото време, вторият фактор, който влияе върху причината за петна между различните видове tRNA: дори ако е необходимо да се разпознае типа кожа и да се прикрепи към новата единична аминокиселина, която ще го покаже. Очевидно е, че стойностите на разпознаването могат да бъдат buti hoch и възпоменателни, но не са необходими, така че работата по разпознаването на гледката на tRNA се трансформира в процес на бижута. В същото време е най-вероятно да се използват аминоацил-тРНК-синтетази: кожата от тях може да се свърже само с една от 20 аминокиселини и те също могат да добавят към тези видове тРНК, като аминокиселини. От таблицата с генетичния код се вижда, че кожната аминокиселина е кодирана от десетична последователност от нуклеотиди, така че например всички тРНК с антикодони CGA, CGG, CGU и CGC ще бъдат разпознати от едни и същи синтетаза, която е преди тях аланин. Такива процеси с една tRNA синтетаза се наричат спорен.
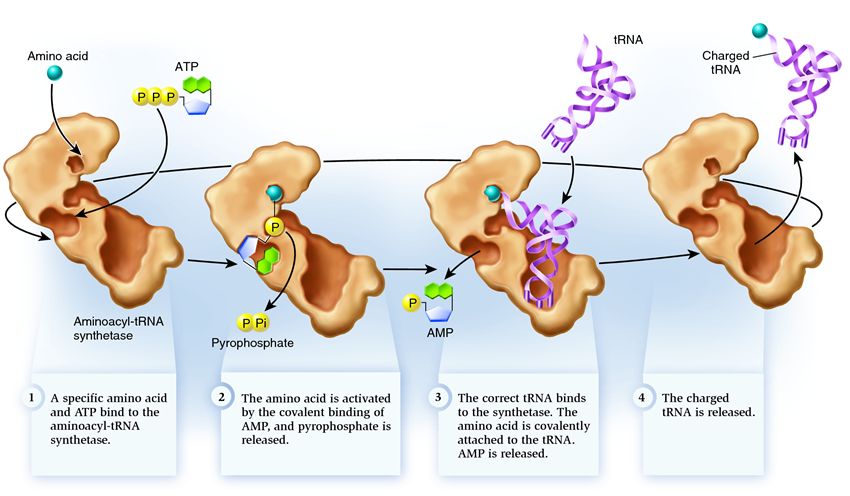
Синтетазата принадлежи към група ензими, чиято функция е свързана със свързването с други молекули и комбинирането им в едно:
1 . синтетазата консумира аминокиселина и ATP молекула. Под формата на АТФ се въвеждат две фосфатни групи, генериращи енергия, необходима за нападателни действия. Аденозин монофосфатът (AMP), който остава под формата на разрушена молекула, се свързва с аминокиселината, подготвяйки я да се свърже с акцепторната фибичка.
2 . синтетазата взема една от различните вериги от аминокиселини на спорадните тРНК.

На този етап е необходима повторна проверка на вида на транспортната РНК синтетаза. Isnuê kílka методи за разпознаване, а в кожната синтетаза е тяхната уникална комбинация. Взаимодействията между тРНК синтетазата включват поне един нуклеотид към антикодон. Акцепторната фиба също зависи от обръщането: тя показва наличието на специфични нуклеотиди, които са от съществено значение за необходимите аминокиселини на спорадични тРНК. Нуклеотидите на други части на tRNA също могат да участват в трансформацията на гените, един по един от основните места на синтетазата. Неправилната tRNA може да расте според необходимостта от някои параметри, но чрез неточен цикъл може да стигне до синтетаза правилно и леко, лесно разпадаща се. И правилната тРНК се прилепва към синтетазата бързо и лесно, след което структурата на синтетазата се променя, стартирайки процеса аминоацилиране за добавяне на аминокиселини към тРНК

3 . аминоацилирането действа върху заместването на смесената AMP молекула с аминокиселината на tRNA молекула. След замяната на AMP, синтетазата се изчерпва и tRNA се подрязва за останалата част от преобразуването на аминокиселината. Ако добавената аминокиселина бъде разпозната като неправилна, тя ще бъде разпозната като tRNA, аминокиселината липсва в синтетазата на спорите и друга молекула може да бъде добавена там. Новата аминокиселина е преминала през етапа на преобразуване на АТФ и тРНК и може да се наложи да бъде преобразувана. Въпреки че помилванията не бяха разрешени, тРНК е заредена с аминокиселина: тя е готова да изиграе своята роля в транслацията на протеини. И синтетазата е готова да приеме нови аминокиселини и тРНК и цикълът ще започне наново.
Ако работата на аминоацил-тРНК-синтетазата е правилна, тя е твърде богата, за да се отложи: ако на този етап ще има грешка, тогава грешната аминокиселина ще бъде добавена към тРНК. Той ще бъде въведен в протеина, който расте върху рибозомата, дори tRNA и рибозомата не функционира като кодон и обръщане на аминокиселини. Последствията от помилването могат да бъдат незначителни или катастрофални, а в хода на естествения подбор с ензими, които не засягат функциите на такива обрати, те са по-важни, което може да предизвика различни варианти за установяване на коректността между аминокиселина и тРНК. Следователно в днешните клетки синтетазата се свързва с грешна аминокиселина средно в една капка от 50 хиляди, а с лека tRNA повече от веднъж на 100 хиляди дни.

Деяките аминокисли се отделят един от един повече от един брой атоми. Просто погледнете техните схеми, става очевидно, че възможността за объркване на аргинин с аланин е много по-малка или погрешно изолевцин с левцин или валин. Следователно кожната синтетаза, която се свързва с едно към едно подобни аминокиселини, може да има допълнителни механизми за обръщане. Оста е пример за такова прикрепване в изолевцин-тРНК синтетаза:

Използва се кожна синтетаза синтетичен център, В който аминокиселините се добавят към тРНК Акцепторната фиба на тРНК, уловена от синтетазата, я консумира сама, като аминокиселина, е готова да влезе в реакция от нея. Работата на тези синтетази ще бъде прекратена, след като аминокиселините и тРНК бъдат произведени. Ale-tRNA-синтетазата може да насърчи ефекта на пардоните чрез използването на други аминокиселини, подобни на изолевцина. Това е всичко корекционен център: Назовете ролята на вината в процеса на производство на тРНК и аминокиселини. Малката фигура вдясно показва, че позицията на края на фибичката на акцепторната тРНК в синтетичния център на Ile-tRNA синтетазата поражда неестествената фибичка. Тим не е по-малко, синтетазата утриму тРНК в тази позиция, докато аминокиселината дойде до нея. Освен това, както се случи, необходимостта от поставяне на акцепторната фиби в синтетичния център е изчерпана и тРНК се изправя, консумирайки върха си с коригиращия център, засаден върху новата аминокиселина.

Очевидно синтетичният център също играе своята роля в усвояването на аминокиселини, които не са подходящи за синтетазата. За да го изразходвате до последно, молекулите трябва да се съобразят с редица умове, кремът на майката е постоянно разширяване. Независимо от тези, че левцинът и изолевцинът отмъщават на един и същ брой атоми, левцинът е по-голям поради присъствието си в обемната структура. Следователно виното не може да проникне в синтетичния център, но изглежда, че е възможно за компактен изолевцин и е лесно да се види Ile-tRNA синтетаза.
И оста на валин, която е най-важната от тези три молекули с подобна атомна структура, лесно заема пространството на изолевцина в синтетичния център и синтетазата идва до тРНК. Самият по този начин да влезе в центъра за групова корекция на синтетазата. Като акцепторна фиба, която се изправя, зарежда се правилно и носи изолевцин, тогава тя не може да се промъкне до центъра на коригиращия център: тя просто е твърде малка за размера на молекулата. В този ред тРНК, която се е изправила, вече не подрязва нищо и ще излезе от синтетазата. И оттук към тРНК се добавя валин, вената преминава в корекционния център и тРНК се получава от синтетазата с нея. Такъв трансцендентален тривал на tRNA повторно свързване се използва за синтетаза с сигнал за помилване, променяйки неговата пространствена конфигурация. В резултат на което:
=) валинът се освобождава от тРНК и се отстранява от синтетазата
=) акцепторният щифт се върти към синтетичния център, пречупвайки се към аминокиселината
=) синтетазата се появява с нова аминокиселина, "зарежда" нейната tRNA и отново проверява, че това е същият изолевцин.
Такъв механизъм на подкожно разпознаване се имплантира от други синтетази.