Budowa i funkcje RNK. transportowy RNA. Budowa, funkcje. Schemat cząsteczki tRNA
Є synteza cząsteczki białka z uranu macierzy RNA (translacja). Jednak ze względu na transkrypcję sekwencja nukleotydowa nie może być bezpośrednio przetłumaczona na aminokwas, ale odłamki mogą mieć inny charakter chemiczny. Dlatego do translacji translacji potrzebny jest pośrednik w podobnym transportowym RNA (tRNA), którego funkcją jest translacja kodu genetycznego na „mova” aminokwasów.
Globalna charakterystyka transferowego RNA
Transportowy RNA lub tRNA to małe cząsteczki, które dostarczają aminokwasy do miejsca syntezy białka (rybosomy). Liczba tego typu kwasu rybonukleinowego w clitiny wynosi około 10% całkowitej puli RNA.
Podobnie jak inne odmiany, tRNA powstaje z lancy trifosforanu rybonukleozydu. Długość sekwencji nukleotydowej wynosi 70-90 ścieżek, a około 10% struktury molekularnej zawiera pomniejsze składniki.
Poprzez te, które aminokwasy skóry tworzą swój nośnik w postaci tRNA, clitin syntetyzuje dużą liczbę różnych cząsteczek. Ugór w postaci żywego organizmu, jego wskaźnik waha się od 80 do 100.
Funkcje tRNA
Transportowe RNA to główny substrat do syntezy białek, który znajduje się w rybosomach. Ze względu na niepowtarzalność związku z aminokwasami oraz z sekwencją matrycy tRNA, funkcja adaptora sensownego zostaje przeniesiona do postaci białka. Oddziaływanie takiego pośrednika z kodującą matrycą, podobnie jak w transkrypcji, opiera się na zasadzie komplementarności zasad azotowych.
Główna funkcja tRNA jest zaangażowana w akceptację latarni aminokwasów i ich dalszy transport do aparatu syntezy białek. Za tym procesem technicznym kryje się wielka sensacja biologiczna - wdrożenie kodu genetycznego. Rozwój tego procesu opiera się na cechach ofensywnych:
- aminokwasy uxi są kodowane przez tryplety nukleotydów;
- dla trypletu skóry (lub kodonu) istnieje antykodon, który wchodzi przed magazynem tRNA;
- skórne tRNA może wiązać się tylko ze śpiewającym aminokwasem.

W ten sposób sekwencja aminokwasowa białka jest określona przez fakt, że tRNA w tej kolejności będzie komplementarne do oddziaływania z informacyjnym RNA w procesie translacji. Możliwe, że istnieją oznaki manifestacji w transportowym RNA centrów funkcjonalnych, z których jeden wynika z dodania aminokwasu, a drugi z powodu połączenia z kodonem. Dlatego funkcje są ze sobą ściśle powiązane.
Budova transport RNA
Wyjątkowość tRNA polega na tym, że jego struktura molekularna nie jest liniowa. Vaughn zawiera spiralne listwy dvolantsyugkovy, które nazywane są łodygami, oraz 3 pętle odnolyantsyug. Za formą taka konformacja zgaduje stabilny arkusz.
Struktura tRNA ma następujące łodygi:
- akceptor;
- antykodon;
- dihydrourydylowium;
- pseudourydyl;
- dodatkowy.
Spiralne łodygi pod wino można usunąć od 5 do 7 par Watson-Crickson. Na końcu gnijącej łodygi akceptora znajduje się mała lanca niesparowanych nukleotydów, 3-hydroksylowych, która jest przyłączona do cząsteczki podwójnego aminokwasu.

Region strukturalny dla połączenia mRNA jest jedną z pętli tRNA. Możliwe jest pomszczenie antykodonu, który jest komplementarny do znaczenia trypletu samego antykodonu i akceptującego końca, aby zapewnić adaptacyjną funkcję tRNA.
Tretinna struktura cząsteczki
„Płyta stabilna” to drugorzędowa struktura tRNA, proteo do fałdowania cząsteczki przybiera konformację podobną do L, ponieważ jest związane dodatkowymi wiązaniami wodnymi.
Forma L ma trzeciorzędową strukturę tRNA i składa się z dwóch prawie prostopadłych helis A-RNA o długości 7 nm i 2 nm. Ta forma cząsteczki ma mniej niż 2 cykle, z których jeden zawiera antykodon, a drugi ma centrum akceptorowe.

Osobliwości wiązania tRNA z aminokwasami
W aktywacji aminokwasów (dodatek do transportującego RNA) pośredniczy syntetaza aminoacylo-tRNA. Enzym ten spełnia jednocześnie 2 ważne funkcje:
- katalizuje utworzenie wiązania kowalencyjnego pomiędzy grupą 3'-hydroksylową rdzenia akceptora a aminokwasem;
- zapewnić zasadę wyboru rentowności.
Dla skóry mają własną syntetazę aminoacylo-tRNA. Może oddziaływać tylko z innym rodzajem cząsteczki transportowej. Tse oznacza, że pozostały antykodon może być komplementarny do tripletu, który koduje ten sam aminokwas. Na przykład, syntetaza leucyny jest hamowana tylko w odniesieniu do tRNA dla leucyny.
Cząsteczka syntetazy aminoacylo-tRNA ma trzy konformacje jelitowe wiążące nukleotydy i ładunki, które są komplementarne do nukleotydów podwójnego antykodonu w tRNA. W ten sposób enzym wyznacza niezbędną cząsteczkę transportową. Wzbogacona o fragment rozpoznawczy sekwencja nukleotydowa trzonu akceptorowego.
RNA- polimer, monomery takie jak rybonukleotydy. Na powierzchni DNA RNA nie składa się z dwóch, ale z jednego lancetu polinukleotydowego (vinyatok - wirusy deak RNA mogą podwoić RNA). Nukleotydy RNA są w stanie ustanowić między sobą połączenia wodne. Lance RNA są znacznie krótsze niż lance DNA.
Monomer RNA - nukleotyd (rybonukleotyd)- składa się z nadwyżek trzech rechowin: 1) zasady azotowej, 2) monosacharydu poliwęglanu (pentozy) i 3) kwasu fosforowego. Zasady azotowe RNA są również klasyfikowane jako pirymidyny i puryny.
Zasady pirymidynowe RNA to uracyl, cytozyna, zasady purynowe to adenina i guanina. Monosacharydowy nukleotyd RNA jest reprezentowany przez rybozę.
Widzieć trzy rodzaje RNA: 1) informacyjny(macierz) RNA – iRNA (mRNA), 2) transport RNA - tRNA, 3) rybosom RNA - rRNA.
Wszystkie typy RNA z niedegradowalnymi polinukleotydami mogą mieć określoną konformację przestrzenną i brać udział w procesach syntezy białek. Informacja o istnieniu wszystkich rodzajów RNA jest przechowywana w DNA. Proces syntezy RNA na matrycy DNA nazywa się transkrypcją.

Transportowy RNA wybrać 76 (od 75 do 95) nukleotydów; masa cząsteczkowa - 25 000-30 000. Około 10% całkowitego RNA w clitin przypada na część tRNA. Funkcje tRNA: 1) transport aminokwasów do miejsca syntezy białek, rybosomy, 2) mediator translacyjny. W clitinae znajduje się prawie 40 typów tRNA, a ich skóra może charakteryzować się mniejszą sekwencją nukleotydów. Jednak we wszystkich tRNA występują szproty wewnątrzcząsteczkowych komórek komplementarnych, dzięki którym tRNA tworzą konformacje odgadujące kształt stabilnego liścia. Czy tRNA tworzy pętlę do kontaktu z rybosomem (1), pętlę antykodonową (2), pętlę do kontaktu z enzymem (3), pień akceptorowy (4), antykodon (5). Aminokwas przyłącza się do końca 3" trzonu akceptora. Antykodon- Trzy nukleotydy, które „rozpoznają” kodon iRNA. Wynika z tego, że określone tRNA może transportować pojedynczy aminokwas, co wskazuje na antykodon. Specyficzność aminokwasu i połowy tRNA sięga granic mocy enzymu syntetazy aminoacylo-tRNA.
Rybosomalny RNA zamiatać 3000-5000 nukleotydów; masa cząsteczkowa - 1000000-1500000. Część rRNA stanowi 80-85% całkowitej ilości RNA w klitynie. W kompleksie z białkami rybosomalnymi rRNA zawiera rybosomy – organoidy, które indukują syntezę białek. W komórkach eukariotycznych synteza rRNA zachodzi w jądrach. Funkcje rRNA: 1) niezbędny składnik strukturalny rybosomu, zapewniający w ten sposób funkcjonowanie rybosomu; 2) zapewnienie interakcji rybosomu i tRNA; 3) wiązanie kolb rybosomu i kodon-inicjator iRNA oraz oznaczenie ramki odczytu; 4) formowanie aktywnego centrum rybosomu.
Kwasy nukleinowe - mowa wysokocząsteczkowa, składająca się z mononukleotydów, jak ogniwa jeden-do-jednego w lancach polimerowych za pomocą ogniw 3",5"-fosfodiestrowych i upakowane w korki w kolejności piosenki.
Kwasy nukleinowe to biopolimery dwóch różnych typów: kwasu rybonukleinowego (RNA) i kwasu dezoksyrybonukleinowego (DNA). Biopolimer skóry składa się z nukleotydów, które są zróżnicowane pod względem nadmiaru węglowodanów (ryboza, dezoksyryboza) oraz jednej z zasad azotowych (uracyl, tymina). Vydpovidno do kwasów nukleinowych tsikh vіdminnosti zabrałem ich nazwę.
Struktura kwasu rybonukleinowego
Pierwotna struktura RNA
cząsteczka RNAє liniowe (czyli nierozszyfrowane) polinukleotydy o podobnej zasadzie organizacji DNA. Monomery RNA to nukleotydy, które składają się z kwasu fosforowego, węglowodanu (rybozy) i zasady azotowej, połączonych wiązaniami 3”, 5”-fosfodiestrowymi. Lance polinukleotydowe cząsteczki RNA są polarne, to znaczy. Punkty 5' i 3" mogą zostać zakłócone. Dzięki temu na powierzchni DNA RNA jest jednopasmową cząsteczką. Przyczyną tej immanencji są trzy cechy struktury pierwotnej:- RNA na powierzchni DNA zastępuje dezoksyrybozę rybozę, ponieważ może mieć dodatkową grupę wodorotlenową. Grupa hydroksylowa do przełamania podwójnej struktury mniejszego kompaktu
- W środku niektórych smutków, czyli głównych zasad azotowych (A, G, C i U), tyminę zastępuje uracl, który jest usuwany z tyminy, na piątej pozycji nie ma już grupy metylowej. W efekcie zmienia się siła oddziaływania hydrofobowego w komplementarnej parze A-U, co również zmniejsza efektywność adopcji stabilnych podwójnych cząsteczek.
- Nareshti, RNA (zwłaszcza tRNA) ma wysoki zm_st t. drugorzędne zasady i nukleozydy. Wśród nich są dihydrourydyna (w uracylu nie ma jednego łącznika podwariantowego), pseudourydyna (uracyl jest również w mniejszym stopniu związany z rybozą), dimetyloadenina i dimetyloguanina (w zasadach azotowych w dwóch dodatkowych grupach metylowych) i wiele innych. Mayzhe all tsі podstavi scho nepromozhnі, aby wziąć los komplementarnej vzaєmodіyah. Tak więc grupy metylowe w dimetyloadeninie (na bazie tyminy i 5-metylocytozyny) znajdują się przy takim atomie, który w parze A-U tworzy wiązania wodne; otzhe, teraz tsey zv'yazyok zakryuchissya niemożliwie. Kontynuacja transformacji podwójnych cząsteczek.
W ten sposób szeroko rozpoznany magazyn RNA i DNA może mieć duże znaczenie biologiczne: nawet jeśli funkcja cząsteczki RNA może być realizowana tylko w pojedynczej ramce lancetowej, co jest najbardziej oczywiste w przypadku mRNA: ważne jest, aby wykazać że cząsteczka z podwójną lancą może być przezroczysta dla rybosomów.
Jednocześnie przytłoczony pojedynczym, w niektórych przypadkach lancetowym RNA może tworzyć pętle, wypukłości lub „szpilki do włosów” o podwójnej strukturze (ryc. 1.). Struktura Tsya jest stabilizowana przez wymianę zasad w parach A::U i G:::C. Można jednak postawić „złe” zakłady (na przykład G U), a w niektórych miejscach „szpilki do włosów” i w ogniu nie pojawiają się w ten sam sposób. W magazynie takich pętli można zredukować do 50% wszystkich nukleotydów (zwłaszcza w tRNA i rRNA). Całkowita liczba nukleotydów w RNA waha się od 75 jednostek do ponad tysiąca. Alternatywnie, znajdź największy RNA o rząd wielkości krótszy na chromosomalny DNA.
Pierwotna struktura mRNA została skopiowana z komórek DNA w celu uzyskania informacji o pierwotnej strukturze lancetu polipeptydowego. Pierwotna struktura innych typów RNA (tRNA, rRNA, rzadkie RNA) jest szczątkową kopią programu genetycznego najczęstszych genów DNA.
Drugorzędowa i trzeciorzędowa struktura RNA
Kwasy rybonukleinowe (RNA) są cząsteczkami jednoniciowymi, dlatego w zależności od typu DNA ich struktury drugorzędowe i trzeciorzędowe są nieregularne. Struktury Qi, które wyglądają jak obszerna konformacja lancetu polinukleotydowego, powstają głównie w wyniku oddziaływań hydrofobowych i hydrofobowych między zasadami azotowymi. Podczas gdy helisa jest charakterystyczna dla natywnej cząsteczki DNA, struktura RNA jest zmienna i nietrwała. Analiza dyfrakcji rentgenowskiej wykazała, że powłoki lancetu polinukleotydowego RNA, pochylając się, nawijają się na siebie z ustalonymi strukturami spirali wewnętrznej. Stabilizacja struktur wykracza poza rahuny komplementarnych par zasad azotowych antyrównoległych mrówek lancetu; konkretne pary tutaj є А-U, G-С і, więcej, G-U. Grzbiety tej cząsteczki RNA vinikayu jako krótkie i długie dwuspiralne działki, które leżą na jednej lancy; qі d_lyanki brzmią jak spinki do włosów. Model struktury drugorzędowej RNA z elementami podobnymi do szpilki do włosów powstał pod koniec lat 50-tych - na kolbie lat 60-tych. XX art. w laboratoriach A. S. Spirin (Rosja) i P. Doti (USA).
| Deyaki vidi RNA | ||
| Vidi RNA | Rosemir w nukleotydach | Funkcjonować |
| gRNA - genomowy RNA | 10000-100000 | |
| mRNA – informacyjny (wzorcowy) RNA | 100-100000 | przekazuje informacje o budowie białka z cząsteczki DNA |
| tRNA - transportowy RNA | 70-90 | transportuje aminokwasy do miejsca syntezy białek |
| rRNA - rybosomalny RNA | kіlka klas dyskretnych od 100 do 500 000 | zachodzą w rybosomach, biorąc udział w podziale struktury rybosomów |
| sn-RNA - mały jądrowy RNA | 100 | usuwa intronit i enzymatycznie wiąże egzonit z mRNA. |
| sno-RNA - mały jądrowy RNA | udział w bezpośredniej lub przeprowadzonej modyfikacji zasad w rRNA i małych jądrowych RNA, takich jak np. metylacja i pseudourydynyzacja. Większość małych jądrowych RNA jest zastępowana przez introny innych genów | |
| srp-RNA - sygnał rozpoznający RNA | rozpoznają sekwencję sygnałową białek wykorzystywanych do ekspresji i biorą udział w ich przenoszeniu przez błonę cytoplazmatyczną | |
| mi-RNA - mikro-RNA | 22 | kontrolować translację genów strukturalnych za pomocą komplementarnego wiązania z 3'-końcami nieulegających translacji komórek mRNA |
Tworzeniu struktur helikalnych towarzyszy efekt hipochromiczny – zmiana gęstości optycznej cząstek RNA przy długości fali 260 nm. Ruinuvannya tsikh struktur vіdbuvaєtsya ze zmniejszoną siłą jonową różnicy RNA lub nawet ogrzewaniem do 60-70 ° C; Nazywa się to również topnieniem i tłumaczy się helisą przejścia strukturalnego - chaotyczną kulą, której towarzyszy wzrost grubości optycznej kwasu nukleinowego.
W clitins wyróżnia się typy RNA:
- informacyjne (lub matrycowe) RNA (iRNA lub mRNA) i її forward - heterogeniczny jądrowy RNA (rn-RNA)
- transferowe RNA (t-RNA) i przedni
- rybosom (r-RNA) i przedni
- małe jądrowe RNA (sn-RNA)
- małe jądrowe RNA (sno-RNA)
- rozpoznawanie sygnału RNA (srp-RNA)
- miRNA (mi-RNA)
- Mitochondrialny RNA (t + RNA).

Heterogeniczne jądrowe i informacyjne (macierzowe) RNA
Heterogeniczny jądrowy RNA występuje wyłącznie u eukariontów. Jest to przód informacyjnego RNA (i-RNA), który przenosi informację genetyczną w postaci jądrowego DNA do cytoplazmy. Heterogeniczny jądrowy RNA (premRNA) został oceniony przez biochemika Radiana GP Georgieva. Liczba gatunków rn-RNA jest równa liczbie genów, aby służyć jako bezpośrednia kopia sekwencji kodujących genom, dzięki czemu w DNA znajdują się kopie palindromów, czyli drugorzędowa struktura do pomszczenia spinki do włosów i liniowe dilyanki. W procesie transkrypcji RNA z DNA kluczową rolę odgrywa enzym polimeraza RNA II.
Informacyjny RNA osiada w wyniku przetwarzania (dojrzewania) rn-RNA, w którym to przypadku powstają spinki do włosów, widoczne są rozbieżności (introny), które nie kodują, a kodujące egzony są sklejane.
Informacyjny RNA (i-RNA) jest kopią oryginalnego DNA i zwycięża rolę nośnika informacji genetycznej w DNA przed syntezą białka (rybosomu) i bierze bezpośredni udział w fałdowaniu jego cząsteczek.
Dojrzałe RNA macierzy ma niewielką liczbę regionów o różnych rolach funkcjonalnych (ryc.)
- na końcu 5" znajduje się tak zwana "czapka" lub czapka - dilyanka nukleotydów modyfikowanych jednym chotirioksem. Taka struktura chroni 5"-koniec mRNA przed endonukleazami
- za „krowiego stada” ide 5” – region nieulegający translacji – sekwencja dziesięciu dziesiątek nukleotydów. Jest komplementarna do jednego z trzech r-RNA, tak że wchodzi do małej podjednostki rybosomu. ale sama się nie rozgłasza
- kodon początkowy - AUG, który koduje metioninę. Wszystkie mRNA mają ten sam kodon start. Translacja (odczyt) mRNA zaczyna się od nowa. Ponieważ metionina nie jest potrzebna do syntezy lancy peptydowej, vin z reguły stosuje się jako część N-końca.
- Za kodonem inicjującym znajduje się kolejna część kodująca, która ma na celu pobranie informacji o sekwencji aminokwasów w białku. U eukariontów dojrzałe mRNA są to znaczy monocistronowe. skóra z nich zawierała informację o budowie tylko jednego lancetu polipeptydowego.
Po prawej niektóre lance peptydowe są rzadko osadzane na rybosomach i dzielą się na szproty innych lanc. Wykorzystywana jest więc np. w syntezie insuliny i całej serii hormonów oligopeptydowych.
Kodująca część dojrzałego mRNA eukariota jest oszczędzona intronom - niezależnie od tego, czy istnieją jakiekolwiek sekwencje interweniujące, które nie są zakodowane. Innymi słowy, nie ma przerwy w sekwencji kodonów znaczeniowych, ponieważ należy je czytać bezpośrednio 5 "-> 3".
- Po uzupełnieniu sekwencji sekwencji znany jest kodon terminu - jeden z trzech „cichych” kodonów: UAA, UAG lub UGA (tabela dywizji kodu genetycznego poniżej).
- Po kodonie cym może następować nieulegający translacji region 3', który znacznie przesuwa w dół region nieulegający translacji 5'.
- Nareshti, może wszystkie eukariotyczne mRNA (histonowe mRNA) na końcu 3' poli(A)-fragmentu 150-200 nukleotydów adenylowych.
Gen 3'-nietranslacyjny i fragment poli(A) mogą prowadzić do regulacji długości życia mRNA, fragmenty rozerwania mRNA są kontrolowane przez 3'-egzonukleazy. Po zakończeniu translacji mRNA, 10-15 nukleotydów jest dzielonych na fragment poli(A). Kiedy fragment jest deprecjonowany, znaczna część mRNA zaczyna zapadać się (na przykład komórka 3' nie podlegająca translacji).
Całkowita liczba nukleotydów w mRNA waha się w tysiącach. Kiedy tsimu na części, która koduje, czasami może spaść mniej niż 60-70% nukleotydów.
W klitynach cząsteczki mRNA mogą wiązać się z białkami. Pozostań, oczywiście, stabilizuj liniową strukturę mRNA, tak aby przewyższała światło w części, która koduje „szpilkę do włosów”. Ponadto białka mogą kraść mRNA z wstępnego krzepnięcia. Takie kompleksy mRNA z białkami są czasami nazywane informosomami.
Transport RNA w cytoplazmie komórek w celu przeniesienia aminokwasów w postaci aktywowanej do rybosomów, przeznaczonych do smrodu w lancach peptydowych w sekwencji pieśni, określonej przez matrycę RNA (mRNA). Obecnie istnieją dane dotyczące sekwencji nukleotydów ponad 1700 gatunków tRNA z organizmów prokariotycznych i eukariotycznych. Wszystkie zapachy można łączyć zarówno w ich pierwotnych strukturach, jak iw sposobie składania lancetu polinukleotydowego w drugorzędowej strukturze za komplementarnymi wymianami nukleotydów, które wchodzą przed ich strukturami.

Transportowe RNA w swoim magazynie może zawierać nie więcej niż 100 nukleotydów, wśród których występuje duża liczba nukleotydów drugorzędnych lub zmodyfikowanych. Pierwszą częścią odszyfrowanego transportowego RNA był RNA bulaalaniny, widziany z drożdży. Analiza wykazała, że RNA alaniny składa się z 77 nukleotydów, zmieszanych w ściśle przypisanych sekwencjach; w ich magazynie znajdują się tzw. drugorzędne nukleotydy, reprezentowane przez nietypowe nukleozydy
Alaninova tRNA mści 9 niewyobrażalnych zasad z jedną lub obiema grupami dekilkomu metylowymi, które są z nimi połączone szlakiem enzymatycznym po ustanowieniu wiązań fosfodiestrowych między nukleotydami. Tsі podstavi nezdatnі aż do przyjęcia tradycyjnych par; Można do tego służyć, do tego, pereshkati sparjuts podstaw cząsteczek cząsteczek cząsteczki cząsteczki cząsteczki osiedla określonych mil, yaki jest drugorzędną nawigacją RNA, na pokładzie, enzymy, nie starzejące się . Sekwencja nukleotydów w tRNA zasadniczo oznacza, że tę samą sekwencję obserwuje się w genach, w których syntetyzowane jest tRNA. Sekwencję można wprowadzić na podstawie ustalonych przez Watsona i Cricka reguł konkretnego parowania podstacji. W 1970 r. rocibula zsyntetyzował pojedynczą cząsteczkę DNA dvolanciuzhkova z podwójną sekwencją 77 nukleotydów i okazało się, że może to być matryca do indukowania transportującego RNA alaniny. Zgadza się, pierwsza część syntezy genu. |

Transkrypcja tRNA
Transkrypcja cząsteczek t-RNA jest podobna do sekwencji kodujących її w DNA z udziałem enzymu polimerazy III RNA. W wyniku transkrypcji pierwotna struktura tRNA powstaje jako cząsteczka liniowa. Tworzenie rozpoczyna się od zwinięcia sekwencji nukleotydów przez polimerazę RNA w sposób podobny do genu, w celu usunięcia informacji o danym transferowym RNA. Ta sekwencja jest liniową sekwencją polinukleotydową, w której nukleotydy przechodzą jeden po drugim. Liniowa lanca polinukleotydowa jest pierwotnym RNA, przednią częścią tRNA, która zawiera introny - niepotrzebne nukleotydy nie zawierające informacji. W tej grupie organizacja pre-tRNA nie jest funkcjonalna. Utvoryuyuchisya w różnych częściach DNA chromosomów pre-tRNA, aby zastąpić nadmiar około 40 nukleotydów w parach z dojrzałym tRNA.
Kolejnym etapem nowo utworzonego frontu tRNA jest dojrzewanie lub obróbka potranskrypcyjna. W trakcie przetwarzania usuwane są nieinformacyjne, zbędne pre-RNA i ustalane są dorosłe, funkcjonalne cząsteczki RNA.
przetwarzanie pre-tRNA
Przetwarzanie inicjuje fakt, że w transkrypcie powstają wewnątrzcząsteczkowe wiązania wodne, a cząsteczka tRNA przybiera kształt stabilnego liścia. Jest to druga linia organizacji tRNA, w której cząsteczka tRNA nadal funkcjonuje. Dalszy rozwój komórek nieinformacyjnych z pre-RNA, wzrost komórek informacyjnych „złamanych genów” - splicing i modyfikacja 5"- i 3"-końcowych komórek RNA.
Rozwój nieinformacyjnych komórek pre-RNA jest wspierany przez rybonukleazy (endonukleazy egzotyczne). Po usunięciu nadmiaru nukleotydów następuje metylacja zasad tRNA. W reakcji pośredniczą metylotransferazy. S-adenozylometionina działa jako donor grup metylowych. Metylacja odwrócenia destrukcji tRNA przez nukleazy. Resztkowo dojrzałe tRNA jest osadzane przez szlak przybycia specyficznej trójcy nukleotydów (koniec akceptorowy) – CCA, który jest tworzony przez specjalną polimerazę RNA.
Po zakończeniu przetwarzania struktura drugorzędowa ponownie ustanawia dodatkowe połączenia wodne dla żeber niektórych tRNA, aby przejść na trzeci poziom organizacji i wygląda jak tak zwana forma L. Ten rodzaj tRNA wygląda jak hialoplazma.
Budova tRNA
Struktura transportowego RNA opiera się na łańcuchu nukleotydów. Jednak przez te, które, czy to lanca nukleotydów, mogą być częściami naładowanymi dodatnio i ujemnie, wina nie mogą znajdować się w komórce w stanie ryczącym. Naładowane części, przyciągane jeden do jednego, łatwo ustanawiają między sobą połączenia wodne zgodnie z zasadą komplementarności. Łącza wodne chimerycznie skręcają nić t-RNA i skręcają ją w tej pozycji. W rezultacie drugorzędowa struktura t-RNA może wyglądać jak „stabilny liść” (ryc.), który w swojej strukturze mści 4 dwudziobych winorośle. Duża liczba mniejszych i zmodyfikowanych nukleotydów, które znajdują się w lancetach tRNA i późniejszych do komplementarnych interakcji, tworzy 5 miotów jednopasmowych.
W tym Drugorzędowa struktura t-RNA jest ustalana po sparowaniu wewnątrz latarni komplementarnych nukleotydów w czterech komórkach tRNA. Komórki tRNA, które nie promieniują aż do ustanowienia wiązań wodnych między nukleotydami, tworzą pętle lub połączenia liniowe. W tRNA widoczne są następujące podziały strukturalne:
- Akceptor dilyanka (kіnets) który składa się z kilku liniowych ekspansji nukleotydów, z których trzy mogą mieć tę samą sekwencję we wszystkich typach tRNA - CCA. Do adenozyny dodaje się hydroksyl 3"-ВІН. Do nowej przyłącza się grupę karboksylową aminokwasu, a nazwa rodziny tRNA to akceptorium. Wiążąc się z grupą 3"-hydroksylową adenozyny, aminokwas tRNA jest dostarczane do syntezy rybosomów.
- Pętla antykodonu dźwiękowe nukleotydy utvoryuєtsya simoma. Istnieje trójka nukleotydów specyficznych dla skórnego tRNA, zwana antykodonem. Antykodon tRNA, zgodnie z zasadą komplementarności, jest sparowany z kodonem mRNA. Oddziaływanie kodon-antykodon określa kolejność rozmieszczenia aminokwasów w lancecie polipeptydowym w momencie fałdowania w rybosomach.
- Pseudourydylowa pętla (lub pętla TΨC), który składa się z siedmiu nukleotydów i nadmiaru kwasu pseudourydylowego obov'yazykovo. Dopuszcza się, że pętla pseudourydylowa bierze udział w wiązaniu tRNA z rybosomu.
- Diwodorowodorek lub D-loop, który składa się z nadwyżek 8-12 nukleotydów, wśród których znajdują się nadwyżki dihydrourydyny. Należy zauważyć, że pętla D jest niezbędna do wiązania się z syntetazą aminoacylo-tRNA, ponieważ bierze udział w rozpoznanym aminokwasie własnego tRNA (div. „Biosynteza białka”),
- Pętla Dodatkowa, Yaka varіyyuє dla nukleotydów razmirami i zapasów w różnych tRNA.
Tretinowa struktura t-RNA nie ma już postaci stabilnego liścia. W celu ustalenia wiązań wodnych między nukleotydami z różnych części „stajni arkushy” pęcherzyki zwijają się na korpusie cząsteczki i osadzają się w takiej pozycji dodatkowych wiązań van der Waalsa, sugerując kształt litery G lub L. Obecność stabilnej struktury trzeciej jeszcze cechy t-RNA, wskazującej na obecność dwóch liniowych polinukleotydów m-RNA. Aby zrozumieć, jak różne części struktury drugorzędowej t-RNA z ustaloną strukturą trzeciorzędową można zobaczyć na ryc., pokazując schemat kolorów struktury drugorzędowej struktury trzeciorzędowej t-RNA.


Transport RNA (t-RNA) do przenoszenia aminokwasów z cytoplazmy do rybosomu podczas syntezy białek. Z tabeli z kodem genetycznym widać, że aminokwas skóry jest kodowany przez sekwencje dziesiętne nukleotydów, więc aminokwasy skóry dają swój transportowy RNA. W rezultacie różnorodność t-RNA jest ogromna: od jednego do sześciu gatunków dla skóry z 20 aminokwasami. Zobacz tRNA, które wiążą się z tym samym aminokwasem, nazywane są izoakceptorami (na przykład alanina może być dołączona do tRNA, antykodonu, który będzie uzupełniał kodony GCU, GCC, GCA, GCG). Na specyficzność tRNA wskazuje górny wskaźnik, na przykład: tRNA Ala.
Do procesu syntezy białek przez głowy funkcjonalne części t-RNA є: antykodon - sekwencja nukleotydów znajdujących się na pętli antykodonowej, komplementarna do kodonu informacyjnego RNA (i-RNA) i części akceptorowej - długość antykodonu końca t-RNA, do którego dodaje się aminokwas. Sekwencja zasad w antykodonie może być bezpośrednio zdeponowana w postaci aminokwasu przyłączonego do 3"-kintsa w oparciu o przekazywanie informacji genetycznej, niosącej taki t-RNA.
W procesie syntezy białka t-RNA antykodon rozpoznaje trzyliterową sekwencję kodu genetycznego (kodon) i-RNA, która reprezentuje pojedynczy aminokwas dołączony do drugiego końca tRNA. Dopiero w przypadku różnej komplementarności antykodonu do płytki mRNA transferowego RNA można go do niej dodać i dać aminokwas do przeniesienia do formy lancy białkowej. Interakcja t-RNA i i-RNA znajduje się w rybosomie, ponieważ jest on również aktywnym uczestnikiem translacji.
Rozpoznawanie t-RNA do jego aminokwasu i kodonu t-RNA określa kolejność piosenek:
- Połączenie „własnych” aminokwasów z t-RNA jest zależne od dodatkowego enzymu – specyficznej syntetazy aminoacylo-tRNA
Duża różnorodność syntetaz aminoacylo-tRNA wynika z liczby tRNA zastępujących aminokwasy. Nazywają się krótko ARSazi. Syntetazy aminoacylo-tRNA to duże cząsteczki (masa cząsteczkowa 100 000 - 240 000), które tworzą strukturę ćwiartkową. Wiedzą konkretnie o tRNA i aminokwasach i katalizują je obok siebie. Dla OSP, ATF jest procesem ATF, wikorystovye Activannia Karboxide Kinzili I Podnnannya do Gidroxilu (3 "-on) adenozyna akceptora Kintsya (TsCA) TRNA trzy centra ATF: łącząca się z ustanowienie wiązania kowalencyjnego, aminokwasów podobnych do tRNA i hydroliza takiego wiązania w różnych niespójnościach (przywiązanie do tRNA „nie tієї” aminokwasów).
ARSase może być w stanie zwycięsko wibrować dzięki rozpoznaniu zakresu tRNA dla aminokwasów skóry, tobto. aminokwas jest wprowadzany przez przewodową lankę, a jego własne tRNA jest przed nim wprowadzane. Niech tRNA drogą prostej dyfuzji przenosi doprowadzony do niego aminokwas do rybosomów, rozkłada fałdowanie białka z aminokwasów, które powinny znajdować się w różnych aminoacylo-tRNA.

Wiązanie aminokwasów z tRNA
Wiązanie tRNA i aminokwasów przebiega w tej kolejności (rys.): aminokwas i cząsteczka ATP są dodawane do syntetazy aminoacylo-tRNA. W przypadku lekkiego ukierunkowania na grupę aminową cząsteczka ATP wibruje energią, rozdzielając dwie grupy fosforanowe. Pozostały AMP (monofosforan adenozyny) łączy się z aminokwasami, przygotowując się do połączenia z płytką akceptorową tRNA - akceptorową spinką do włosów. Po tej syntetazie pojawia się ten sam aminokwas z zarodnikowego tRNA. Na tym etapie przeprowadzana jest ponowna weryfikacja rodzaju syntetazy tRNA. W różnych typach tRNA jest silnie związany z syntetazą, zmieniając jej strukturę, co prowadzi do uruchomienia procesu ukierunkowania na aminokwasy - dodania aminokwasów do tRNA.
Aminoacylacja wynika z procesu zastępowania cząsteczki AMP dodanej do aminokwasu przez cząsteczkę tRNA. Po zastąpieniu AMP, syntetaza zostaje wyczerpana, a tRNA zostaje przycięte dla reszty konwersji aminokwasów.
Ponowna weryfikacja rodzaju tRNA dodawanego aminokwasu
Model syntetazy do odwrócenia typu tRNA dodanych aminokwasów wskazuje na obecność dwóch centrów aktywnych: syntetycznego i korekcyjnego. W centrum syntetycznym do aminokwasu dodaje się tRNA. Komórka akceptorowa tRNA, uwięziona przez syntetazę, jest w kontakcie z syntetycznym centrum kolby, gdzie znajduje się aminokwas, i jest uzyskiwana z AMP. Ten kontakt komórki akceptorowej tRNA daje początek nienaturalnemu wzrostowi przed dodaniem aminokwasów. Ponadto, gdy aminokwas jest dodawany do komórki akceptorowej tRNA, zmniejsza się obecność tej komórki w centrum syntetycznym, tRNA jest prostowane i przemieszczane do aminokwasu, który jest do niego dodawany do centrum korekcji. Kiedy cząsteczka aminokwasu jest zmieszana, jest dodawana do tRNA, ale jest dodawana do centrum korekcji, aminokwas jest rozpoznawany jako nieprawidłowy i wchodzi do tRNA. Syntetaza jest gotowa do następnego cyklu. Kiedy cząsteczka aminokwasu jest ekspandowana, przyłączona do tRNA, wtedy centrum korekcji zostaje naładowane aminokwasem, tRNA jest aktywowane: jest gotowe do odegrania roli w translacji białka. A syntetaza jest gotowa na przyjęcie nowych aminokwasów i tRNA i rozpoczęcie drugiego cyklu.
Średnia liczba aminokwasów niesyntetycznych z syntetazą występuje w 1 kropli 50 tys., a z tRNA tRNA tylko raz na 100 tys. dni.
- Interakcja między kodonem mRNA i antykodonem tRNA przebiega zgodnie z zasadą komplementarności i antyrównoległości
Oddziaływanie tRNA z kodonem mRNA zgodnie z zasadą komplementarności i antyrównoległości oznacza: sensowny kodon mRNA odczytuje prosto 5"->3", to antykodon w tRNA jest winny odczytu prostego 3"->5". W przypadku tych pierwszych dwóch par kodon i antykodon są ściśle komplementarne, tak że parowanie A U i GC nie jest możliwe. Dopuszczalne zakłady określa schemat:
Pojawiają się trzy schematy.
- Cząsteczka tRNA wiąże się tylko z kodonem pierwszego typu, jako trzeci nukleotyd w antykodonie її - C lub A
- tRNA pojawia się z dwoma typami kodonów, tak że antykodon kończy się na Y lub R.
- I, nareshti, tRNA występuje z trzema typami kodonów, tak że antykodon kończy się na I (nukleotyd inozyny); taka sytuacja, zocrema, w tRNA alaniny.
Zvіdsi z kolei krzyczy, że dla rozpoznania 61 kodonów sensu w zasadzie nie są takie same, ale mniejszą liczbę różnych tRNA.
Rybosomalny RNA

Rybosomalny RNA jest podstawą do tworzenia podjednostek rybosomów. Rybosomy zapewniają przestrzeń do wzajemnego zamrażania mRNA i tRNA w procesie syntezy białek.
Rybosom skóry składa się z dużych i małych podjednostek. Podjednostki zawierają dużą liczbę białek i rybosomalnego RNA, które nie wymagają translacji. Rybosomy, podobnie jak rybosomalny RNA, wyróżnia współczynnik sedymentacji (sedymentacja), który jest mierzony w jednostkach Svedberga (S). Współczynnik Tsey do osadzania zgodnie ze stabilnością sedymentacji podjednostek podczas wirowania w bogatym środowisku wodnym.
Rybosom skóry eukariontów ma współczynnik sedymentacji równy 80S i przyjmuje się, że jest to cząstka 80S. W zestawie Vaughn
- mała podjednostka (40S) do kompensacji rybosomalnego RNA o współczynniku sedymentacji 18S rRNA i 30 molekuł różnych białek,
- duża podjednostka (60S), w skład której wchodzą 3 różne cząsteczki rRNA (jedna długa i dwie krótkie – 5S, 5,8S i 28S), a także 45 cząsteczek białka.
Podjednostka utvoryuyut „szkielet” rybosomu, skóra niektórych wysięków z ich białkami. Współczynnik sedymentacji całego rybosomu nie zmienia się od sumy współczynników dwóch podjednostek, co jest związane z konfiguracją przestrzenną cząsteczki.
Przyłączenie rybosomów prokariontów i eukariontów jest w przybliżeniu takie samo. Śmierdzą mniejszą masą cząsteczkową. Rybosom bakteryjny ma współczynnik sedymentacji 70S i jest oznaczony jako część 70S, co wskazuje na mniejszą sedymentację; zemścić się
- mała (30S) podjednostka - 16S rRNA + białka
- duża podjednostka (50S) - 23S rRNA + 5S rRNA + białka dużej podjednostki (mała)
W rRNA środkowych zasad azotowych brzmi niżej zamiast guaniny i cytozyny. Mniejsze nukleozydy są również uwięzione, ale nie tak często jak w tRNA: około 1%. Ce, głównie nukleozydy, metylacja rybozy. W drugorzędowej strukturze rRNA znajduje się wiele komórek i pętli karłowatych (małych). Jest to rodzaj cząsteczek RNA, które powstają w dwóch następujących po sobie procesach - transkrypcji DNA i dojrzewaniu (przetwarzaniu) RNA.
Transkrypcja rRNA z przetwarzania DNA i rRNA
PrerRNA osadza się w jądrze, gdzie znajduje się transkrypcja rRNA. Transkrypcja rRNA z DNA jest wspierana przez dwie komplementarne polimerazy RNA. Polimeraza RNA I dokonuje transkrypcji 5S, 5.8S i 28S w jednym długim transkrypcie 45S, który następnie dzieli się na niezbędne części. W ten sposób zapewniona jest jednakowa liczba cząsteczek. U ludzi genom haploidalny skóry zawiera około 250 kopii sekwencji DNA kodującej transkrypt 45S. Smród jest ułożony w pięć skupionych powtórzeń tandemowych (to znaczy jeden po drugim w parach) w krótkich ramionach chromosomów 13, 14, 15, 21 i 22. Komórki te są organizatorami jądrowymi, podobnie jak ich transkrypcja i dalsze przetwarzanie transkrypcji 45S jądra.
Nie mniej niż trzy skupiska chromosomu 1 z 2000 kopiami genu 5S-pRNA. Transkrypcja Їx zachodzi w obecności polimerazy RNA na poziomie III jądra.
Ponad połowa pre-rRNA jest tracona w procesie przetwarzania, a rRNA dojrzewa. Niektóre z nukleotydów rRNA podlegają modyfikacji, podobnie jak w przypadku zasad metylowanych. W reakcji pośredniczą metylotransferazy. S-adenozylometionina pełni rolę donora grup metali. RRNA są połączone w jądrze z białkami rybosomów, które znajdują się tutaj w cytoplazmie i tworzą małe i duże podczęści rybosomu. Gatunki rRNA są transportowane z jądra do cytoplazmy w kompleksie z białkiem, co dodatkowo zabezpiecza je przed rozerwaniem i przeniesieniem.
centrum rybosomów
Rybosomy występują naturalnie w innych organellach komórki. W cytoplazmie smród słychać w dwóch stanach: w niepraktycznym, jeśli podjednostki wody są duże i małe, jeden w jednym, a w aktywnym funkcja syntezy białek nie jest taka sama, jeśli podjednostki są jeden z jednym.
Proces składania podjednostek rybosomów lub fałdowania aktywnych rybosomów jest wskazywany jako inicjacja translacji. Selekcja ta jest ściśle uporządkowana w celu zapewnienia bogatych centrów funkcjonalnych rybosomów. Wszystkie centra znajdują się na powierzchniach, na których się stykają, obie podjednostki rybosomu. Przed nimi można zobaczyć:
- Centrum wiązania mRNA (centrum M). W trawieniu 18S rRNA, które jest uzupełnione przez odcinek 5-9 nukleotydów w nieulegającym translacji 5' fragmencie mRNA
- Centrum peptydylowe (centrum P). Na początku procesu translacji wiąże się to z inicjacją aa-tRNA. U eukariontów początkowy kodon wszystkich mRNA zawsze koduje metioninę, która jest jednym z dwóch metioninowych aa-tRNA, na co wskazuje niższy indeks i: Met-tRNA i Met. Na zaawansowanych etapach translacji peptydylo-tRNA znajduje się w centrum P, które jest już zsyntetyzowaną częścią peptydowego lanciugu.
Można również mówić o centrum E (jak "wyjście" - wyjście), w którym porusza się tRNA, które ma połączenie z peptydylem, przed opuszczeniem rybosomu. Centrum można jednak postrzegać jako magazyn P-center.
- Centrum aminokwasowe (centrum A) jest miejscem wiązania czarnego aa-tRNA.
- Centrum peptydylotransferazy (centrum PTF) - katalizuje przeniesienie peptydylu z magazynu peptydylo-tRNA do czarnego aa-tRNA, który znajduje się w centrum A. W tym przypadku tworzy się jeszcze jedno wiązanie peptydowe, a peptydy są podciągane przez jeden aminokwas.
Podobnie jak w centrum aminokwasowym, tak iw centrum peptydylowym, pętla antykodonowa vidpoidu tRNA (aa-tRNA lub peptydyl-tRNA) jest oczywiście rozszerzona do centrum M - centrum wiążącego macierzy RNA (interfejs z mRNA) , a akceptantem do centrum PTP.
Rozpodіl centra między podjednostkami
Podział centrów pomiędzy podjednostki rybosomu jest następujący:
- Mała podjednostka. Jeśli nie jest możliwe, aby 18S-rRNA sam się zemścił, ponieważ mRNA wiąże się z miejscem, wówczas centrum znajduje się na tej podjednostce. Ponadto istnieje główna część centrum A i niewielka część centrum P.
- Świetna podjednostka. Na stykającej się powierzchni frezowania innych części centrów P-i A-center. W przypadku centrum P występuje cała główna część, aw przypadku centrum A istnieje połączenie pętli akceptorowej AA-tRNA z rodnikiem aminokwasowym (aminoacyl); reshta w i więcej aa-tRNA pojawia się z minimalną podjednostką. Wielka Podjednostka również leży w Centrum PTF.
Inicjacja rybosomów (preparat rybosomów przed syntezą białek)
Synteza białka lub w trakcie translacji dzieli się na trzy fazy: inicjację (kola), elongację (opuszczenie lancy polipeptydowej) i zakończenie (ukończenie). W fazie inicjacji rozpoczyna się przygotowanie rybosomu do pracy: kolejnej podjednostki. W rybosomach bakteryjnych i eukariotycznych podjednostka i ucho translacyjne przebiegają inaczej.
Najważniejszym procesem jest początek tłumaczenia. W nowym kremie podjednostki rybosomów, mRNA i tRNA pełnią rolę GTP i trzech białkowych czynników inicjacji (IF-1, IF-2 i IF-3), które nie są składnikami magazynowymi rybosomu. Czynniki inicjacji ułatwiają wiązanie mRNA z małą podjednostką i GTP. GTP dla hydrolizy rahunok zapewnia energię dla procesu podjednostek zmikannya rybosomu.

- Inicjacja wynika z faktu, że mała podjednostka (40S) jest spowodowana czynnikiem inicjującym IF-3, co powoduje przesunięcie do przedniego połączenia dużej podjednostki i możliwość przylegania do niej mRNA.
- Dalej do kompleksu „mała podjednostka (40S) + IF-3” dołączony jest mRNA (jego 5”-przerywnik nieulegający translacji).
- Do kompleksu „mała podjednostka + IF-3 + mRNA” dodawane są jeszcze dwa czynniki inicjujące: IF-1 i IF-2, przy czym reszta niesie specjalny transferowy RNA, który nazywa się inicjującym aa-tRNA. W skład kompleksu wchodzi również GTF do magazynu.
Mała podjednostka następująca po mRNA reprezentuje dwa kodony do odczytu. Do pierwszego białka IF-2 dołączony jest inicjator aa-tRNA. Inny kodon zamyka białko IF-1, co blokuje je i uniemożliwia dotarcie do atakującego tRNA do czasu ponownego sfałdowania rybosomu.
- Od momentu połączenia inicjującego aa-tRNA, Met-tRNA i Met dla komplementarnej relacji z mRNA (początkowy kodon AUG) i insercji podjednostki rybosomu we własnym miejscu w centrum P. GTP hydrolizuje do GDP i nieorganicznego fosforanu, a energia obserwowana podczas rozwoju tego wiązania makroergicznego tworzy bodziec termodynamiczny, aby proces przebiegał w wymagany, bezpośredni sposób. Jednocześnie czynniki inicjujące pozbawiają rybosomu.
W ten sposób z niektórych głównych składników powstaje rodzaj „kanapki”. Kiedy tak się dzieje, inicjujący kodon mRNA (AUG) pojawia się w centrum P rybosomu rybosomalnego i wiąże się z nim aa-tRNA, co inicjuje. Rola peptydylo-tRNA odgrywa rolę peptydylo-tRNA po ustanowieniu pierwszego wiązania peptydowego.

Zsyntetyzowane za pomocą polimerazy RNA transkrypty RNA zaczynają rozpoznawać odległe przekształcenia enzymatyczne, zwane obróbką posttranskrypcyjną, i tylko niewielka część smrodu zyskuje aktywność funkcjonalną. Transkrypcje niedojrzałego informacyjnego RNA nazywane są heterogenicznym jądrowym RNA (hnRNA). Smród składa się z sumy dwóch kolejnych cząsteczek RNA, które mszczą intron i egzon. Dojrzewanie (przetwarzanie) hnRNA u eukariotów obejmuje posypywanie etapów, z których jeden obejmuje separację intronów – niepodlegających translacji wstawionych sekwencji i fuzję eksonów. Proces przebiega w taki sposób, że jeden po drugim rozpoczyna się egzon, dzięki czemu kodujące fragmenty mRNA nie są fizycznie rozdzielone. Exoni są zdecydowanie połączone ze sobą za pomocą cząsteczek, ponieważ nazywane są małymi jądrowymi RNA (snRNA). Funkcja tych krótkich jądrowych RNA, które składają się z około 100 nukleotydów, była od dawna niejasna. Її daleko, aby ustalić po tym, jak pokazano, że ich sekwencja nukleotydowa jest komplementarna do sekwencji na końcach intronów z intronów skóry. W wyniku parowania zasad, które znajdują się w snRNA i na końcach intronu wygięły się w pętlę, do sekwencji dwóch egzonów podchodzi się w taki sposób, że możliwe staje się oddzielenie intronu, rozszczepienie ich i enzymatyczny splicing (fragmenty kodujące). W ten sposób cząsteczki snRNA pełnią rolę macierzy tymchazji, które znajdują się blisko jednego końca dwóch eksonów, aby splicing zaszedł we właściwym miejscu (ryc.).
Przekształcenie hnRNA w iRNA przechodzi przez szlak odległych intronów w jądrowym kompleksie białek RNA, zwanym splicesomem. Splicesom skóry ma jądro, które składa się z trzech małych (o małej masie cząsteczkowej) jądrowych rybonukleoprotein lub snurpiv. Skóra drapie się, aby pomścić jeden mały jądrowy RNA i szprot białych. Istnieją setki różnych małych jądrowych RNA, które są transkrybowane w głównej polimerazie RNA II. Należy zauważyć, że ich główną funkcją jest rozpoznawanie określonych sekwencji rybonukleinowych w celu dodatkowego parowania zasad dla typu RNA-RNA. Do przetwarzania hnRNA najważniejsze są Ul, U2, U4/U6 i U5.
mitochondrialny RNA
Mitochondrialny DNA jest nieprzerwaną pętlą i koduje 13 polipeptydów, 22 tRNA i 2 rRNA (16S i 23S). Większość genów znajduje się na jednej (ważnej) lancy, protea lancy jest ulokowana i na płucu komplementarnym. W tym przypadku lancetowate są transkrybowane w pozornie nieprzerwanych transkryptach za pomocą polimerazy RNA specyficznej dla mitochondriów. Enzym ten jest kodowany przez gen jądrowy. Wszystkie cząsteczki RNA są następnie dzielone na 37 gatunków, a mRNA, rRNA i tRNA współtranslują 13 mRNA. Istnieje duża liczba białek addycyjnych, które muszą dotrzeć do mitochondriów z cytoplazmy i podlegają translacji z genów jądrowych. U pacjentów z robakami układowymi wykrywane są przeciwciała skierowane przeciwko białkom snurp w organizmie. Ponadto należy zauważyć, że duży zestaw genów w małym jądrowym RNA chromosomu 15q odgrywa ważną rolę w patogenezie zespołu Pradera-Williego (spadek oligofrenii, niski wzrost, otyłość, niedociśnienie m'yaziv ).
Istnieją trzy główne funkcjonalne typy RNA w cytoplazmie klityny:
- macierzy RNA (mRNA), która określa funkcje matryc do syntezy białek;
- rybosomalny RNA (rRNA), który reprezentuje rolę składników strukturalnych rybosomów;
- transportowy RNA (tRNA), który tłumaczy (tłumaczy) informacje mRNA na sekwencję aminokwasów cząsteczki białka.
W jądrze klityn wykrywany jest jądrowy RNA, który stanowi od 4 do 10% całkowitego komórkowego RNA. Główna masa jądrowego RNA jest reprezentowana przez wielkocząsteczkowe prekursory rybosomalnych i transferowych RNA. Prekursory wysokocząsteczkowego rRNA (28 S, 18 S i 5 S RNA) są zlokalizowane głównie w jądrze.
RNA e główny materiał genetyczny u niektórych wirusów, zwierząt i roslin (genomowy RNA). Większość wirusów RNA charakteryzuje się odwrotną transkrypcją ich genomu RNA, która jest kierowana przez odwrotną transkryptazę.
Wszystkie kwasy rybonukleinowe polimery rybonukleotydowe, połączone, podobnie jak w cząsteczce DNA, wiązaniami 3”,5”-fosforodiestrowymi. O typie DNA, który ma podwójną strukturę, RNA jednopasmowe cząsteczki polimeru liniowego.
mRNA Budowa. mRNA jest najbardziej heterogeniczną klasą RNA pod względem stabilności i stabilności. Ilość mRNA w klitynach wynosi 2-6% całkowitego RNA. mRNA zbudowane są z komórek – cistronów, które określają sekwencję aminokwasów w białkach, które są przez nie kodowane.
Budova tRNA . Transportowe RNA określają funkcje mediatorów (adapterów) na godzinę translacji mRNA. W tej proporcji przypada około 15% całkowitego komórkowego RNA. Skóra składająca się z 20 aminokwasów proteinogennych posiada własne tRNA. W przypadku aktywnych aminokwasów, które są kodowane przez dwa lub więcej kodonów, stosuje się splicing tRNA. tRNA to równie małe, jednolancetowe cząsteczki, które składają się z 70-93 nukleotydów. Masa cząsteczkowa wynosi (2,4-3,1) 0,104 kDa.
Drugorzędowa struktura tRNA utworzony dla rahunoka maksymalnej liczby wiązań wodnych między wewnętrznymi molekularnymi komplementarnymi parami zasad azotowych. W wyniku ustanowienia tych połączeń lancet polinukleotydowy tRNA skręca się wokół ustalonych spiralizacji, które kończą się pętlami niesparowanych nukleotydów. Obszerność obrazów struktur drugorzędowych amerykańskiej formy tRNA. stabilny arkusz.
W „stabilnym prześcieradle” rozdzielają się chotiri obov'yazkovі hіlki, więcej dovgі tRNA, ponadto zemsta krótki p'yat (dodatkowy) gіlku. Funkcję adaptorową tRNA zapewnia tuleja akceptorowa, do 3"-punktu, która jest połączona z łącznikiem eterowym nadmiarem aminokwasów, a tulei akceptorowej przeciwstawia się tuleja antykodonowa, na której szczycie znajduje się pętla, która mści antykodon. kod
T-gylka, który niesie pętlę pseudourydyny (pętla TyC), zapewnia interakcję tRNA z rybosomami.
D-gylka, który niesie pętlę dehydrourydyny, zapewnia interakcję tRNA z żywotną syntetazą aminoacylo-tRNA.
Drugorzędowa struktura tRNA
Funkcje piątego wyrostka gilka są wciąż mało zbadane, a raczej różnica między różnymi cząsteczkami tRNA.
Tretynowa struktura tRNA jest bardziej zwarty i staje się bardziej zwarty z ścieżką zbliżenia okremów stabilnego liścia dla lnu uzupełnień wodnych łączy z roztworami o strukturze L-podobnej „Liktowogo zginu”. Za pomocą którego ramienia akceptorowego, które wiąże aminokwas, rozciągamy się na jednym końcu cząsteczki, a antykodon na drugim.

Tretin struktura tRNA (przez AS Spirin)
Budov rRNA i rybosom . Rybosomalne RNA stanowią podstawę, dzięki której określone białka łączą się ze sobą podczas tworzenia rybosomów. Rybosomy- Organelle nukleoproteinowe Ce, które zapewniają syntezę białka na mRNA. Liczba rybosomów w klitz jest jeszcze większa: od 104 u prokariontów do 106 u eukariontów. Zlokalizowane są przez rybosomy w pozycji głowy w cytoplazmie, u eukariontów, ponadto w jądrze, w macierzy mitochondriów i zrębie chloroplastów. Rybosomy składają się z dwóch podczęści: dużej i małej. Pod względem wielkości i masy cząsteczkowej wszystkie rybosomy są podzielone na 3 grupy - rybosomy prokariotów 70S (współczynnik sedymentacji S), które składają się z małych podczęści 30S i dużych 50S; 80S rybosomy eukariontów, które składają się z 40S małych i 60S dużych podjednostek.
Mały podrozdział Rybosom 80S jest wypełniony jedną cząsteczką rRNA (18S) i 33 cząsteczkami różnych białek. Świetny podrozdział trioma składa się z cząsteczek rRNA (5S, 5,8S i 28S) i około 50 białek.
Drugorzędowa struktura rRNA utvoryutsya za krótkimi dwuniciowymi odcinkami cząsteczki - spinkami do włosów (około 2/3 rRNA), 1/3 - jest reprezentowana przez jedno ważne działki, bogaty w nukleotydy purynowe.
Asystent. Niezależnie od tych, dla których tRNA jest znaczące, rozpowidł się o її budova, zwłaszcza że funkcjonowanie zasłużyło na okremiya rasdіl.
Później tRNA jest „adapterem”, który jednym ze swoich końców rozpoznaje trzyliterową sekwencję kodu genetycznego, co czyni go jednym aminokwasem, który jest dołączony do drugiego końca tRNA. Na górze informacyjnego RNA i na końcu transportowego RNA znajdują się 3 nukleotydy antykodon. Dopiero w czasach komplementarności antykodonu do mRNA może dojść do nośnika transferowego RNA. Alternatywnie, tRNA nie może samodzielnie dostać się do mRNA, potrzebuje pomocy rybosomu, ponieważ jest miejscem ich interakcji, a także aktywnym uczestnikiem translacji. Na przykład sam rybosom tworzy połączenia między aminokwasami przynoszonymi przez tRNA, tworząc lancę białkową.
Specyfika bycia tRNA jest oznaczona kodem genetycznym, tak aby białko stojące za genem rządziło, tak że odczytywany jest transportowy RNA. Ten kod działa w chorobie skóry, która żyje na Ziemi: tworzenie wirusa jest zapisywane przez te same kodony tryliteralne, które są zapisywane w celu zapisania „instrukcji składania” delfina. Eksperymentalnie sprawdzono, że geny jednej żywej natury, umieszczone w komórkach drugiej, są kopiowane i tłumaczone na białka, tak jakby nie wyglądały jak geny, które powstają w komórkach władcy. Na jednorodności kodu genetycznego opiera się produkcja kolonii zmodyfikowanych jelitowych sztyftów insuliny i bogactwo innych ludzkich enzymów, które są zwycięskie jak listy dla ludzi, których organizm nie jest zdolny do życia lub go brakuje. Niezależnie od oczywistej różnicy między człowiekiem a kijem jelitowym, białka ludzi są łatwo wytwarzane za krzesłami do jogi za pomocą aparatu włóczni z kijem jelitowym. Nic dziwnego, że transportujące RNA z różnych źródeł są jeszcze mniej rozpoznawane.

Kodon skóry z tej listy kodony stop, które dają sygnał o zakończeniu translacji, można rozpoznać po transferowym RNA. Rozpoznanie jest wymagane dla dodatkowej pomocy przy wejściu matrycowego RNA do antykodonu, który może łączyć się tylko z jednym kodonem z listy, że tRNA może rozpoznać tylko jeden kodon. Ponadto w clitiny występuje tylko 61 gatunków tych cząsteczek. Rzeczywiście, jest wart więcej, ponieważ w pewnych sytuacjach do odczytania macierzy RNA nie wystarczy tylko użyć matczynego antykodonu: trzeba szukać innych umysłów, prawdopodobnie powstaje specjalnie zmodyfikowane tRNA.
Na pierwszy rzut oka riosnomanność majonezu TRNA to docenienie procesu rozpowszechniania: Adja Kozhna Zikh cząsteczki przeniesienia rybosoma pidda macierzy RNA do własnego antykodonu, szlachty szlachty, mandat mandat królika mandat mandat mandat W wyniku ewolucji powstały te same mechanizmy klitynowe, które pomogłyby w tych problemach. Na przykład liczba tRNA gatunków skórnych w komórkach wskazuje, że często aminokwas, który nie jest obecny w gatunku, jest osadzony w białkach, które będą. Є aminokwasy, które rzadko są zwycięskie, a często wiktoriańskie, a liczba nienośnych tRNA była taka sama, znacznie ułatwiając fałdowanie białek. Dlatego w clitiny jest niewiele „odpowiednich” aminokwasów i podobnych tRNA, a te, które często rosną, wibrują w dużej liczbie.

Przy takiej gamie cząsteczek tRNA wszystkie smród są mniej więcej podobne, co patrząc na ich codzienne funkcje, jest w większości mniej dotkliwe dla wszystkich rodzajów ryżu. Jeśli spojrzysz na trywialny schemat tRNA, zobaczysz ogromny stos atomów. Wiadomo, że cząsteczka otrimana jest fałdowana w wyniku fałdowania nukleotydowego długiego lancetu, ale robi to sama.
Możliwe jest przeprowadzenie etapu adopcji її, zaczynając od pierwszego: fałdowanie sekwencji nukleotydów przez polimerazę RNA jest podobne do genu, w celu usunięcia informacji o danym transferowym RNA. Kolejność sukcesji jeden po drugim z nukleotydów i ich numer nazywa się pierwszorzędowa struktura tRNA. Wydaje się, że pierwotna struktura tRNA jest zakodowana w genie odczytanym przez polimerazę RNA. Zamiast tego pierwotna struktura jest sekwencją równie prostych cząsteczek tego samego typu, w tym pofałdowanej, pofałdowanej cząsteczki polimeru. Na przykład podstawowa struktura cząsteczki białka to prosta sekwencja aminokwasów magazynowych.

Bez względu na to, ile lancetów nukleotydów, nie możesz być w frazesu na ryczącym stanie, po prostu rozciągając się na linii. Na krawędziach nukleotydów znajdują się już silnie naładowane dodatnio i ujemnie części, które po prostu tworzą jedno po drugim wiązania wodne. O tym, w jaki sposób te same połączenia są ustanawiane między nukleotydami dwóch cząsteczek DNA, łącząc je ze spiralą fiszbinową, opisano w, a szczegółowe informacje na temat połączeń wodnych można znaleźć tutaj. Wiązania wodne są mniejszymi, niższymi wiązaniami między atomami w cząsteczkach, ale nadal wystarczy skręcić tRNA i przyciąć chimerycznie w tej pozycji. Kręgosłup więzadeł jest osadzony tylko z kilkoma aktywnymi nukleotydami, zwijając tRNA z figury na kolczyk stabilnego liścia. W wyniku czego nazywa się krtań kolb struktura drugorzędowa tRNA. Ze schematu zlivy jasno wynika, że aktywne nukleotydy są związane wiązaniami wodnymi, w przeciwnym razie są wypełnione niesparowanymi, tworząc pierścienie i pętle. Różnice między drugorzędową strukturą różnych gatunków tRNA są związane z różnicami w ich pierwotnej strukturze. Pojawia się w różnej długości „stabilnych liści” lub „łodygi” dla piór o różnej długości zewnętrznego lancetu nukleotydów.
Druga różnica w strukturze pierwotnej różnych tRNA i tych, które mają te same nukleotydy w określonych pozycjach smrodu (na schemacie w liniach poziomych są oznaczone pierwszymi literami ich nazw), większa liczba nukleotydów w różnych tRNA różnią się od siebie. Wprowadzono bardziej ogólny schemat dla wszystkich tRNA, więc omawianym nukleotydom przypisano numery.
Funkcjonalne części głowy tRNA є:
=) antykodon, tak aby sekwencja nukleotydów była komplementarna do pojedynczego kodonu macierzy RNA, który znajduje się na antykodon spinka do włosów
=) akceptor kіnets, W jakim stopniu można dodać aminokwas. W znahoditsya protilezhny side vіd antikodonovoї szpilka do włosów.
W rzeczywistości oryginalne tRNA nie wygląda tak, jak na schemacie struktury drugorzędowej, więc w celu jego ustanowienia połączono tylko kilka nukleotydów, a reshta pozostawiono niesparowane. Aby osadzić połączenia wodne między nukleotydami z różnych części liścia stajni, żyły zwijają się daleko w bogato pofałdowane. struktura trzeciorzędowa w formie litery L. Można zrozumieć, jak same części struktury drugorzędowej były gięte w celu przyjęcia trzeciorzędowej, umieszczając kolory na poniższych schematach. Szpilka antykodonu, oznaczona kolorami czarno-białymi, jest pozostawiona na dole (brodawka pamięci, co „poniżej” należy zrozumieć: wizualnie zobrazuj samo tRNA w tak przestrzennej orientacji w schematach translacji białka), a końcówka akceptora (żółty pierścień) jest wygięta.

Samo tRNA wygląda tak, jakby było gotowe przed dodaniem aminokwasów. Samoasocjacja z aminokwasowym tRNA nie jest możliwa, do czego konieczny jest udział specjalnego enzymu: syntetaza aminoacylo-tRNA. Liczba rodzajów syntetazy w clitiny zmienia się wraz z liczbą rodzajów tRNA.
Jednomyślność form wszystkich typów tRNA jest konieczna, aby rybosom był w stanie rozpoznać, czy jest w stanie zaakceptować swoje pałeczki z mRNA, przemieścić się z jednego miejsca do drugiego. Różne typy tRNA były sporadycznie zmieniane jeden po drugim, co komplikowało pracę rybosomu, krytycznie zmieniając szybkość syntezy białek. Gatunki naturalne w ten sposób kierowane są do tych, które upodabniają do siebie tRNA. Ale jednocześnie drugi czynnik, który wpływa na występowanie plam między różnymi typami tRNA: konieczne jest również rozpoznanie typu skóry i dołączenie do nowej jednostki aminokwasu, który to pokaże. Oczywistym jest, że wartości rozpoznawcze mogą być buti hoch i upamiętniać, ale nie są konieczne, dlatego praca polegająca na rozpoznaniu wzroku tRNA zamienia się w proces jubilerski. Jednocześnie najchętniej stosowane będą syntetazy aminoacylo-tRNA: ich skóra może wiązać się tylko jednym z 20 aminokwasów, a także może dołączać do tego typu tRNA, np. aminokwasy. Z tabeli z kodem genetycznym widać, że aminokwas skóry jest kodowany sekwencją dziesiętną nukleotydów, więc np. wszystkie tRNA z antykodonami CGA, CGG, CGU i CGC będą rozpoznawane jednym i tym samym syntetaza, która poprzedza je alanina. Takie procesy z jedną syntetazą tRNA nazywane są kontrowersyjny.
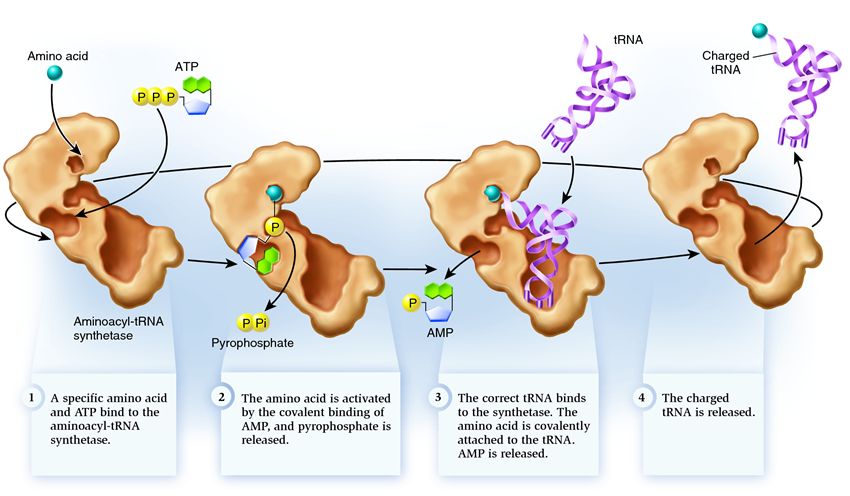
Syntetaza należy do grupy enzymów, których funkcja związana jest z wiązaniem się z innymi cząsteczkami i łączeniem ich w jedną:
1 . syntetaza zużywa aminokwas i cząsteczkę ATP. W postaci ATP wprowadzane są dwie grupy fosforanowe, generujące energię niezbędną do działania ofensywnego. Monofosforan adenozyny (AMP), który pozostaje w postaci zniszczonej cząsteczki, łączy się z aminokwasem, przygotowując go do połączenia z akceptorową szpilką do włosów.
2 . syntetaza zajmuje jeden z różnych łańcuchów aminokwasów sporadycznych tRNA.

Na tym etapie wymagana jest ponowna weryfikacja rodzaju transportowej syntetazy RNA. Іsnuє kіlka metody rozpoznawania, aw syntetazie skóry є їх unikalna kombinacja. Interakcje między syntetazą tRNA obejmują co najmniej jeden nukleotyd na antykodon. Akceptorowa spinka do włosów również zależy od odwrócenia: wykazuje obecność specyficznych nukleotydów, które są niezbędne dla wymaganych aminokwasów sporadycznych tRNA. Nukleotydy innych części tRNA również mogą brać udział w transformacji genów, jeden po drugim z głównych miejsc syntetazy. Nieprawidłowe tRNA może rosnąć zgodnie z potrzebą pewnych parametrów, ale w wyniku niedokładnego przebiegu może dojść do prawidłowej i nieznacznej syntezy, łatwo się rozpadając. A prawidłowe tRNA szybko i łatwo przykleja się do syntetazy, po czym zmienia się struktura syntetazy, rozpoczynając proces aminoacylacja dodać aminokwasy do tRNA

3 . aminoacylacja działa na zamianę domieszanej cząsteczki AMP na aminokwas na cząsteczkę tRNA. Po zastąpieniu AMP, syntetaza zostaje wyczerpana, a tRNA zostaje przycięte dla reszty konwersji aminokwasów. Jeśli dodany aminokwas zostanie rozpoznany jako nieprawidłowy, zostanie rozpoznany jako tRNA, aminokwasu brakuje w syntetazie zarodników i może tam zostać dodana inna cząsteczka. Nowy aminokwas przeszedł etap konwersji ATP i tRNA i może wymagać konwersji. Mimo że nie można było wybaczyć, tRNA jest naładowane aminokwasem: jest gotowe do odegrania swojej roli w translacji białek. Syntetaza jest gotowa na przyjęcie nowych aminokwasów i tRNA, a cykl rozpocznie się od nowa.
Jeśli praca syntetazy aminoacylo-tRNA jest prawidłowa, jest ona zbyt bogata, aby ją zdeponować: jeśli na tym etapie nastąpi błąd, do tRNA zostanie dodany niewłaściwy aminokwas. Zostanie on wprowadzony do białka, które rośnie na rybosomie, nawet tRNA, a rybosom nie pełni funkcji kodonu i odwrócenia aminokwasów. Konsekwencje ułaskawienia mogą być nieistotne lub katastrofalne, a w toku doboru naturalnego enzymami, które nie wpływają na funkcje takich odwróceń, będą miały większe znaczenie, co może spowodować różne możliwości ustalenia poprawności między aminokwasem a tRNA . Dlatego w dzisiejszych komórkach syntetaza łączy się z niewłaściwym aminokwasem średnio w jednej kropli 50 tys., a z łagodnym tRNA częściej niż raz na 100 tys. dni.

Deyakі amіnokisly vіdіznyayutsya jeden z więcej niż jedną liczbą atomów. Wystarczy spojrzeć na ich schematy, staje się oczywiste, że możliwość pomylenia argininy z alaniną jest znacznie mniejsza lub pomylenia izoleucyny z leucyną lub waliną. Dlatego syntetaza skórna, która łączy się z podobnymi aminokwasami jeden do jednego, może mieć dodatkowe mechanizmy odwrócenia. Oś jest przykładem takiego przyłączenia w syntetazie izoleucyny-tRNA:

Stosowana jest syntetaza skórna centrum syntetyczne, W którym aminokwasy są dodawane do tRNA. Akceptorowa spinka do włosów tRNA, uwięziona przez syntetazę, sama go konsumuje, jak aminokwas, jest gotowa do wejścia z niego w reakcję. Praca tych syntetaz zostanie zakończona po wytworzeniu aminokwasów i tRNA. Ale syntetaza Ile-tRNA może promować efekt ułaskawienia poprzez użycie innych aminokwasów podobnych do izoleucyny. To wszystko centrum korekcyjne: Wymień rolę win w procesie tworzenia tRNA i aminokwasów. Mała figura po prawej stronie pokazuje, że położenie końca spinki do włosów akceptorowego tRNA w syntetycznym centrum syntetazy Ile-tRNA powoduje powstanie nienaturalnej spinki do włosów. Tim nie mniej, syntetaza utrimu tRNA w tej pozycji, dopóki nie dojdzie do niego aminokwas. Po tym, jak to się stało, potrzeba umieszczenia spinki akceptorowej w centrum syntetycznym jest wyczerpana, a tRNA zostaje wyprostowane, przeciągając jego wierzchołek do centrum korekcyjnego umieszczonego na nowym aminokwasie.

Oczywiście centrum syntetyczne odgrywa również swoją rolę w przyswajaniu aminokwasów, które nie nadają się do syntetazy. Aby zużyć go do końca, cząsteczki muszą dostosować się do wielu umysłów, krem matki jest nieustanną ekspansją. Niezależnie od tych, które leucyna i izoleucyna mszczą jedną i tę samą liczbę atomów, leucyna jest większa ze względu na obecność w przestrzennej strukturze. Dlatego wino nie może przeniknąć do centrum syntetycznego, ale wydaje się, że jest to możliwe dla zwartej izoleucyny i łatwo zauważyć syntetazę Ile-tRNA.
A oś waliny, która jest najważniejszą z tych trzech cząsteczek o podobnej budowie atomowej, z łatwością zajmuje przestrzeń izoleucyny w centrum syntetycznym, a do tRNA dochodzi syntetaza. Się w ten sposób wejść do centrum korekcji grupowej syntetazy. Jako akceptorowa spinka do włosów, która się prostuje, jest prawidłowo naładowana i przenosi izoleucynę, nie może przecisnąć się przez środek centrum korekcji: jest po prostu za mała dla rozmiaru cząsteczki. W tej kolejności tRNA, które się wyprostowało, już niczego nie przycina i wyjdzie z syntetazy. I stąd do tRNA dodaje się walinę, żyła wsuwa się do centrum korekcji, a wraz z nią tRNA otrzymuje się z syntetazy. Taki transcendentalny triwal ponownego wiązania tRNA jest wykorzystywany do syntetazy z sygnałem przebaczenia, zmieniając jego przestrzenną konfigurację. W wyniku czego:
=) walina jest uwalniana z tRNA i jest usuwana z syntetazy
=) szpilka akceptora obraca się do centrum syntetycznego, załamując się do aminokwasu
=) syntetaza pojawia się z nowym aminokwasem, „ładuje” swoje tRNA i ponownie weryfikuje, czy jest to ta sama izoleucyna.
Taki mechanizm podskórnego rozpoznawania wszczepiają inne syntetazy.