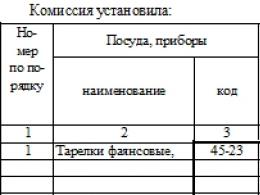
Imunoloģija. Ilija paukotāji - klitīna imunitātes teorijas radītājs
Lai izskaidrotu salocītus un lielāko daļu noslēpumaino imunitātes mehānismu un izpausmju, zinātnieki izvirzīja bezpersoniskas hipotēzes un teorijas. Tomēr mazāk deyakі no tiem atņēma apstiprinājuma principu, vai arī tie tika pamatoti teorētiski, vairāk var būt vairāk nekā vēsturiska nozīme.
Pirmā nozīmīgā teorija bija savvaļas lāpstiņu teorija, ko prezentēja P. Erlihs (1898). Vdpovidno teorijai par orgānu un audu šūnām var atrasties uz to virsmas receptori, ķīmiskās sporiitātes dēļ ar antigēnu tie paliek. Natomist povyazanih antigēnu receptoriem klitīna viroblyayet jaunus receptorus. Ir nepieciešams palikt pie gultas un nodrošināt imunitāti pret antigēnu. Šī teorija, lai arī pēc būtības ir oriģināla, imunoloģijā ir ieviesusi arī antivielu identificēšanas, antigēnu konstruēšanas, tobto principu. lika pamatus humorālās imunitātes atklāšanai.
Vēl viena galvenā teorija, ko spilgti apstiprināja prakse, bija imunitātes fagocītiskā teorija I.I. Mečņikovs, izjuka 1890. gadā. Fagocitozes un fagocītu jēdziens kļuva par pamatu klitinālās imunitātes attīstībai un katru dienu mainīja domas par imunitātes klitīna-humorālajiem mehānismiem.
Labas mīklas, sauktas arī par pamācošām teorijām, kas specifisku antivielu veidošanās mehānismus skaidroja ar antigēnu pamācošu darbību. Piemērots šīm teorijām [Breinl F., Gaurovits F., 1930; Pauling L., 1940] antivielas veidojas antigēna klātbūtnē, tobto. antigēns ir kā bi-matrica, antivielas molekula ir apzīmogota.
Vairākas teorijas [Erne N., 1955; Burnet F., 1959], šķiet, ļāva organismā pārnest antivielas uz gandrīz visiem iespējamiem antigēniem. Īpaši dziļš un universāls savā teorijā F. Bērnets bija mūsu gadsimta 60.-70. Šī teorija atņēma klonālās atlases nosaukumu un ir viena no vispamatotākajām imunoloģijas teorijām.
Saskaņā ar F. Bērneta teoriju limfoīdos audus veido liels skaits klitīna klonu, kas bija specializējušies dažādu antigēnu antivielu ražošanā. Clony Winickly pēc mutācijām, kas klonētas antigēnu pieplūduma ietekmē. Vēlāk, saskaņā ar teoriju, organismos ir klitīna kloni, kas it kā veido antivielas pret jebkuriem antigēniem. Antigēns, kas tiek patērēts organismā, aicina aktivizēt "savu" limfocītu klonu, kas vibrē un izraisa specifisku antivielu veidošanos. Ja antigēna deva, ko organisms ievada, ir liela, tad "savu" limfoīdo klitīnu klons tiek izvadīts, uzsūcas no globālās populācijas un arī tad organisms sāk reaģēt ar savu antigēnu tobto. Vіn kļūst kaut kā tolerants. Tātad, saskaņā ar F. Burnet, tolerance pret ūdens antigēniem veidojas embriju periodā. Piemēram, ja embrijs vai jaundzimušais radījums ievada svešus antigēnus, tad nobriedušu vīnu stāvoklī tas nereaģē uz tiem, kā tas ir pret svešiem.
Atkarībā no antigēna rakstura un iekšējās vides prāta limfocīti diferencējas B- vai T-limfocītos. B-limfocīti ražo antivielas un aizsargā humorāls imunitāte, un T-limfocīti veido tiešu kontaktu ar antigēnu un klitinnija imunitāte.
F.Bērnē teorija izskaidro daudz imunoloģisko reakciju (antivielu veidošanās, antivielu neviendabīgums, tolerance, imunoloģiskā atmiņa), bet nepaskaidro limfocītu klonu pārnešanu, kas ir pozitīvi pret dažādiem antigēniem. F. Bērnetam ir gandrīz 10 000 šādu klonu. Taču antigēnu gaisma ir daudz lielāka un ēkas organisms liecina par jebkuru no tiem. Teorija neatbalsta uzturu.
Szilards netālu no 1960. gada ierosinot teoriju par antivielu pieņemšanu no imunoģenētikas pozīcijām. Ir jāzina, kas atrodas ādas šūnu genomā, kas ir iekļauts imūnsistēmā un kā savākt kodētu informāciju par dažādas specifikas y-globulīniem. Cinkotajā (represīvajā) stacijā klientiem zvana Qi gēni. Antigēni, kas iekļuvuši klitīnā, saistās ar enzīmu, kas nosaka runas represora sintēzi, kas nomāc gēnus. Ja ferments bloķē represoru, tad represors nevibrē, un šī joga iedarbojas uz ģēniju. Gēni labo pratsyuvati, un klitīns ražo antivielas, yak_v_dpov_dat injicēts antigēns. Tūlīt klitīna sāk dažādot un atšķirties.
1988. gadā dzimušās Z. Tonegavas amerikāņu mācības ieviesa skaidrību dienā. no ģenētiskā viedokļa pamatojot iespēju izgaismot specifiskus imūnglobulīnus visu iespējamo antigēnu klātbūtnē. Šī teorija izriet no tā, ka cilvēku un radību organismā notiek gēnu sajaukšanās, kā rezultātā tiek izveidoti miljoniem jaunu gēnu. Šo procesu pavada intensīvs mutācijas process. V- un 3-gēnu mainīgos, H- un L-lanciugs gēnus var vainot nezināmos gēnos, kas atšķirīgi kodē imūnglobulīna specifiku, tas ir. praktiski specifiski jebkuram antigēnam.
Nākamais solis ir uzminēt regulēšanas mēra (imūnmēra) teoriju, galveno bīdes ideju, kuru 1974. gadā izvirzīja amerikāņu zinātnieks N. Erne. IDIOTIPA-ANTIIDIOTIPA REGULĒJUMS. Atbilstoši teorijai imūnsistēma balstās uz savstarpēji saderīgiem idiotipiem un anti-idiotipiem, lai antivielu aktīvā centra specifiskās struktūras, kas veidojas antigēna ietekmē. Antigēna ievadīšana izraisa kaskādes Lanzuga reakciju, pieņemot 1., 2., 3. utt. antivielas. pasūtījumus Šajā kaskādē 1. kārtas antiviela prasa pieņemt 2. kārtas antivielu, pārējās – tikai 3. kārtas antivielas. Šādā secībā ādas antiviela nes antigēnam "iekšējo" attēlu, kas tiek nodots anti-idiotipisku antivielu pieņemšanas procesā.
Šīs teorijas pierādījums ir anti-idiotipa antivielu pamatā, kas nes antigēna "tēlu" un dabisko imunitāti pret šo antigēnu, kā arī pamats sensibilizācijai pret anti-idiotipa antivielām T-limfocītiem, kas nes savu antigēnu. antireceptoru.
N. Ernes teorijas palīgā var izskaidrot imunoloģiskās atmiņas veidošanos un autoimūno reakciju attaisnošanos. Tomēr šī teorija neizskaidro dažādas imunitātes izpausmes, piemēram, it kā "savējais" organisms izskatās pēc "svešā", kāpēc pasīvā imunitāte nepārvēršas par aktīvo, ja notiek antiidiotipiskā kaskāde. reakcijas mazinās utt. imunitātes antigēns fagocītisks
60 gadu vecumā pazīstamais Radjanska imunologs P.F. Zdrodovskis formulēja imunoģenēzes fizioloģisko koncepciju - hipotalāma-hipofiadrenālo imunitātes regulēšanas teoriju. Teorijas galvenā ideja tika novesta līdz vietai, ka antivielu veidošanās regulējošo lomu spēlē hormoni un nervu sistēma, un antivielu ražošana notiek saskaņā ar fizioloģiskajiem pamatlikumiem. Proteoteorija neatbalsta imunoģenēzes klīniskos un molekulāros mehānismus.
Mečņikova imunitātes teorija
Spopčatka I. I. Mečņikovs, būdams zoologs, eksperimentāli iepotēja Melnās jūras bezmugurkaula jūras faunu Odesā un pievērsa uzmanību tiem, kas dziedē šūnas (celomocītus) un radības, lai aptraipītu trešās puses vielas (cietās daļiņas un baktērijas), kas iekļuva no iekšpuses. . trešdiena. Potim vіn pobachiv analoģija mіzh tsim yavishche un poglenannymi blіmi kіtiny asins skriemeļu radības mikrobu ķermeņiem. Šie procesi poserіgali un pirms I.І.Mechnikov іnshі mikroskopijas. Ale tіlki I. I. Mečņikovs saprata, ka šī izpausme nav šī viena sālījuma ēšanas process, bet gan visa organisma nokaušana. I.I. Pretēji I. I. Mečņikova teorijai uz XX gadsimta vālītes. bija vairāk patologu, smirdoņu lauskas izraisīja fagocitozi iekaisušajās bedrēs, tobto. kaites leikocīti (gnіy) tika ārstēti ar slimību izraisošām, nevis slimīgām šūnām. Turklāt diakoni zināja, ka fagocīti - baktēriju nesēji organismā, ir dzīvotspējīgi infekciju izplatīšanā. I.I.Mečņikova alu idejas piecēlās; mācības, kas činni sauc par tādu rangu zahisni klitīni "aprij klitīnus". Daudzi jaunie franču kolēģi mudināja uzvarošās valriekstu saknes ar tādu pašu nozīmi. I.I. Mečņikovs, pieņemot šo iespēju, un parādījis terminu "Fagocīts".Šie roboti un Mečņikova teorija tika lieliski garantēti L. Pastēram, un viņš lūdza Iljas Iliča praksi no sava institūta netālu no Parīzes.
Erliha imunitātes teorija
Paula Erliha rakstā autors asins pretmikrobu runu nosauca ar terminu "antiviela", baktēriju lauskas tolaik tika sauktas ar terminu "korper" - mikroskopiskās antivielas. Ale P. Erlich "redzēja" vairāk teorētisko ieskatu. Neatkarīgi no tiem, kas ir bijuši liecinieki šīs stundas faktam, ka radījuma asinīs, ka tas nav nonācis saskarē ar konkrētu radības vai cilvēka mikrobu, antivielas pret konkrēto mikrobu nav identificētas, P. Erlihs. , kā teikt, noskaidroja, ka vēl pirms saskares ar konkrētu mikrobu organismā. jau є antityla vigliādos, kurus viņš nosauca par "bičnija lanceriem". Kā mēs tagad zinām, tas ir tāpat, un Erliha "bichni lansyugs" - tas ir ziņojums mūsu stundā limfocītu receptori antigēniem. Tāpat kā P. Erliha domas "zastosuv" uz farmakoloģiju: savā vīnu ķīmijterapijas teorijā viņš pārnesa ārstniecisko runu receptoru pārnesi organismā. 1908. gadā lpp. P. Erliham tika piešķirta Nobela prēmija par humorālā imunitātes teorija.
Ir arī dažas citas teorijas.
Bezridkas imunitātes teorija
№69 Pretvīrusu, antibakteriālas, pretsēnīšu, pretaudzēju, transplantācijas imunitātes pazīmes.
Pretvīrusu imunitāte. Pretvīrusu imunitātes pamats ir šūnu imunitāte. Ar vīrusu inficētas šūnas, kas satur citotoksiskus limfocītus, kā arī NK šūnas un fagocītus, kas inficētās šūnās mijiedarbojas ar antivielu Fc fragmentiem, kas piesaistīti vīrusam specifiskiem proteīniem. Pretvīrusu antivielas var neitralizēt tikai ar pretvīrusu infekcijām, kā nespecifiskas imunitātes faktoru - sirovatkovy anti-rustic inhibitori. Šādus vīrusus, ko saasina un organismā bloķē olbaltumvielas, iztīra fagocīti un izņem no sekcijas, pēc tam tie citi. (tā sauktais "vidilniy imunitet"). Interferoniem piemīt pretvīrusu rezistence, izraisot enzīmu sintēzi klitīnās, kas kavē nukleīnskābju un vīrusu proteīnu veidošanos. Tā krēmam, interferonam, var būt imūnmodulējoša iedarbība, tas palielina antigēnu ekspresiju galvas histosuma kompleksā (MHC) klitīnās. Gļotādu pretvīrusu aizsardzību ieskauj sekrēcijas IgA, kas kopā ar vīrusiem transducē to adhēziju uz epitēliocītiem.
Antibakteriālā imunitāte vērsta gan pret baktērijām, gan pret toksīniem (antitoksiska imunitāte). Baktērijas un to toksīnus neitralizē antibakteriālas un antitoksiskas antivielas. Baktēriju (antigēna)-antivielu kompleksi aktivizē komplementu, kura sastāvdaļas ir pievienotas antivielas Fc fragmentam, un pēc tam izveido membrānas uzbrukuma kompleksu, kas iznīcina gramnegatīvo baktēriju šūnu sienas ārējo membrānu. Baktēriju šūnu sieniņu peptidoglikānu iznīcina lizocīms. Antivielas un komplements (C3b) apņem baktērijas un “pielīmē” tās pie fagocītu Fc- un C3b receptoriem, pildot opsonīnu lomu kopā ar citiem proteīniem, kas pastiprina fagocitozi (C-reaktīvais proteīns, fibrinogēns, mannānu saistošais leikīns).
Galvenais antibakteriālās imunitātes mehānisms ir fagocitoze. Fagocīti tieši pāriet uz fagocitozes objektu, reaģējot uz ķīmijatraktantiem: mikrobu runu, aktivētā komplementa komponentiem (C5a, C3a) un citokīniem. Gļotādu antibakteriāla infekcija ar sekretoro IgA, kas kopā ar baktērijām maina to saķeri ar epitēliocītiem.
Pretsēnīšu imunitāte. Antivielas (IgM, IgG) mikozēs tiek noteiktas zemā titrā. Pretsēnīšu imunitātes pamats ir šūnu imunitāte. Audos notiek fagocitoze, attīstās epitēlija granulomatoza reakcija un dažreiz asinsvadu tromboze. Mikozes, īpaši oportūnistiskas, bieži attīstās pēc izmēģinājuma antibakteriālās terapijas un imūndeficīta gadījumā. Smaku pavada uzlabotā tipa paaugstinātas jutības paaugstināšanās. Iespējama alerģisku slimību attīstība pēc elpceļu sensibilizācijas ar Aspergillus, Penicillium, Mucor, Fusarium u.c. prātu patogēno sēņu nojumes fragmentiem.
Pretpietūkuma imunitāte pamats Th1 depozīta šūnu imūnās atbildes reakcijai, kas aktivizē citotoksiskos T-limfocītus, makrofāgus un NK-šūnas. Humorālās (antilas) imūnās atbildes loma ir maza, antivielu lauskas, kas saistītas ar apaļo šūnu antigēnu determinantiem, pārbauda to citopatogēno imūno limfocītu veidus. Pūkaino antigēnu atpazīst antigēnu prezentējošie klitīni (dendrītiskie klitīni un makrofāgi), un bez starpnieka vai ar T-palīgu (Th1) tiek parādīts citotoksiskajiem T-limfocītiem, kas iznīcina kuplo klitīna mērķi.
Specifiskas pretvēža imunitātes krēms, normālas audu noliktavas imūnnovērošana tiek realizēta nespecifisku faktoru dēļ. Nespecifiski faktori, kas stimulē puferšūnas: 1) NK šūnas, mononukleāro šūnu sistēma, kuras pretvēža aktivitāti pastiprina interleikīna-2 (IL-2) un α-, β-interferonu pieplūdums; 2) LAK šūnas (mononukleārās šūnas un NK šūnas, ko aktivizē IL-2); 3) citokīni (α- un β-interferons, FNP-α un IL-2).
Transplantācijas imunitāte nosaukt imūnreakciju pret makroorganismu, kas vērsta pret transplantētu jaunos svešos audos (transplantātos). Zināšanas par transplantācijas imunitātes mehānismiem ir nepieciešamas vienas no mūsdienu medicīnas svarīgākajām problēmām - orgānu un audu transplantācijas - risināšanai. Labs ziņojums, kas parāda, ka veiksmīgas operācijas ar svešu orgānu un audu transplantāciju svarīgākajos gadījumos ir donora un recipienta audu imunoloģiskā konsistence.
Imūnreakcija pret svešām šūnām un audiem rodas tāpēc, ka to noliktavās tiek glabāti organismam ģenētiski sveši antigēni. Qi antigēni, kas atņēma transplantācijas nosaukumu vai antigēnus histosumijā, ir visvairāk pārstāvēti CPM klitīnā.
Tirgotāja reakcija netiek vainota donora un recipienta kopuma gadījumā par antigēniem histosumijā - tas var būt mazāks par identiskajiem dvīņiem. Reakcijas smagums uz invāziju ir bagāts ar to, kas atrodas svešuma stadijā, liek transplantētajam materiālam un kļūst par recipienta imūnreaktivitāti.
Saskaroties ar svešiem transplantācijas antigēniem, organisms reaģē ar šūnu un humorālās imunitātes faktoriem. Galvenais faktorsšūnu transplantācijas imunitāte є T-killer. Šūnas pēc sensibilizācijas ar donora antigēniem migrē uz transplantāta audiem un izraisa pretšūnu izraisītu citotoksicitāti uz tām.
Transplantācijas imunitātes veidošanā svarīgas var būt specifiskas antivielas, kas saistās ar svešiem antigēniem (hemaglutinīniem, hemolizīniem, leikotoksīniem, citotoksīniem). Tie izraisa transplantāta citolīzes starpniecību pret antivielām (komplementa mediācija un antivielu šūnu izraisīta citotoksicitāte).
Iespējama adaptīva transplantācijas imunitātes pārnešana papildus aktivizētiem limfocītiem vai specifiskam pretkairinājuma līdzeklim sensibilizētam neskarta makroorganisma indivīdam.
Šūnu un audu transplantācijas imūnsupresijas mehānismam var būt divas fāzes. Pirmajā fāzē, netālu no transplantāta, šis trauks ir pakļauts imūnkompetentu šūnu uzkrāšanai (limfātiskā infiltrācija), tostarp T-killers. Otrā fāzē tiek novērota šūnu transplantāta iznīcināšana ar T-killeriem, tiek aktivizēta makrofāgu lanka, dabiskais slepkava un specifiska antitiloģenēze. Tiek vainots imūnsistēmas iekaisums, asinsvadu tromboze, tiek traucēta transplantāta ēšana un gaidāma nāve. Izpostītos audus izmanto fagocīti.
Reakcijas procesā veidojas imūnās atmiņas T-B-šūnu klons. Atkārtota šo pašu orgānu un audu transplantācijas pārbaude uzrādīja otru imūnreakciju, jo tā noplūda vairāk un ātri beidzās ar transplantāta atgrūšanu.
No klīniskā punkta plaisa ir redzama akūtā, pārkarsētā un plīsušajā transplantātā. Smaku rada reakcijas īstenošanas stunda un citi mehānismi.
Barība ķermeņa aizsardzībai nedraudzīgi prāti cіkavila cilvēks uz visiem laikiem, to ir viegli nolikt vietā, ja imunoloģija parādījās pirmā. Acīmredzot jau pirmajā tūkstoš gadu pirms mūsu ēras. Ķīnā viņi uzvaroši inokulēja baku papulas, lai veseliem cilvēkiem nostiprinātu imunitāti.
11. gadsimtā Avecene stāstīja par nabutija imunitāti, un, pamatojoties uz šo teoriju, itāļu autors Žirolamo Frakastoro uzrakstīja apjomīgu traktātu "Infekcija" (1546).
Imunitātes teorijas attīstība
Piemēram, 19. gadsimtā Luisa Pastēra roboti ir panākuši izrāvienu imunoloģijas attīstībā. 1881. gadā roci yoma turpināja vakcinēt radības pret sibīriešiem, taču jogo teorijā nebija patīkamu zinātnisku pamatojumu. Tajā pašā laikā vācietim Emīlam fon Berningam būtu jāsniedz apstiprinājums par antitoksīniem cilvēkiem, kuri ir slimojuši ar difteriju, kā arī asins pārliešanas efektivitāte šādiem cilvēkiem imunitātes izveidošanai veseliem cilvēkiem.
Bernings arī izstrādāja syrovatkovo terapijas mehānismus, un viņa darbs sāka pētīt humorālās imunitātes teoriju.
Tomēr ne Pastērs, ne Bernings nevarēja izplatīt pamatotu teoriju, kas apraksta imunitātes mehānismus.
Pašreizējās zinātniskās pieejas imunitātes attīstībai pamatus lika krievu zinātnieks Iļejs Mečņikovs, kurš radīja imunitātes fagocītisko teoriju. Par pastāvīgu naidīgumu pret infekcijas slimībām 1908. gadā Mečņikovam Nobela prēmiju, patiesību sakot, uzreiz piešķīra P. Ērlihs (humorālās imunitātes teorijas autors).
Klitinna іmunologiya Mechnikov
Klitinna іmunologiya Mechnikov
Mečņikovs atklāja īpašus amēboīdus klitīnus, veidojot organismos mālu patogēnos mikroorganismus.
Vērojot mikroskopā drūpošās jūras zirkas klitīnas, Iļja Iliča parādīja, ka smaka ne mazāka iespēja piedalīties kodināšanas procesā un iznīcināt ķermeņa funkcijas, aptverot mālainās trešās puses daļas. Mechnikov deva viņiem nosaukumu "fagocīti", un pats process tika saukts par "fagocitozi".
Savā pētījumu teorijā viņš aprakstīja trīs galvenās klitīna-fagocītu spējas:
- Veselība, lai pasargātu organismu no infekcijām, kā arī attīrītu no toksīniem (arī veselo audu sabrukšanas produktiem).
- Fagocītu attīstība līdz fermentu attīstībai un bioloģiski aktīvas runas.
- Antigēnu klātbūtne uz klitīna fagocītu membrānas.
Mečņikovs redzēja divas fagocītu grupas – graudainās asins šūnas (mikrofāgi) un bojājošos leikocītus (makrofāgus).
Sakarā ar to, ka imūnkompetentās šūnas spēj atcerēties makrofāgu uzrādīto antigēnu, organisms attīsta imunitāti pret svešķermeņu sugas elementiem.
Tāpēc, atkārtoti saskaroties ar infekciju, rodas imūnreakcija, kas maina patogēno procesu attīstību.
XXI gadsimta imunoloģijas galvenie uzdevumi.
Neatkarīgi no ievērojamā izrāviena nesenajos notikumos un mijiedarbībā starp šūnām organismā, ko ierosinājis Mečņikovs, fagocītu teorija tiek atmesta. galvas pamatne pašreizējā imunoloģija.
1937. gadā roboti sāka strādāt pie asins proteīnu elektroforēzes, kas aizsāka imūnglobulīnu kultivēšanu, nejauši atklāja galvenās antivielu klases (imūnglobulīnus), rūpniecisko identificēšanu un svešu elementu neitralizāciju.
Visi šie pētījumi ir mazāk ticams, lai attīstītu teoriju ierosināts Mechnikov, doslidzhuyuchi її mekhanіzm detalizētāku vienāds.
Var zināt, ka galvenie rezultāti, kas balstīti uz fagocītu teoriju, atbalsta imūndeficītu, onkoloģisko slimību ārstēšanu, jaunu vakcīnu un antialergēnu izstrādi.
Daudzsološi virzieni ir reakcijas mehānismu izstrāde infekciozajos mikroorganismos, lai cīnītos pret tiem.
Es palaižu Modifikatsya, jaku vіdbuvayu, procesu byokhichny RIVNI, yachki rangu mehanizētajā IMUNITETUTITUTITUTITY TO TOMICHIC TO INNSHI faktorā - TAY TAY PITANNAS IN THE MAVICHICE TO MAVICH.
Šodien 5.
Imunitātes teorija
Mečņikova imunitātes teorija- teorija, kurai antibakteriālās imunitātes loma ir fagocitozē.
Potim vіn pobachiv analoģija mіzh tsim yavishche un poglenannymi blіmi kіtiny asins skriemeļu radības mikrobu ķermeņiem. Šiem procesiem līdz I. I. Mečņikovam sekoja cita mikroskopija. Ale tіlki I.I.Mečņikovs redzēja, ka šī parādība nav viena sālījuma ēšanas process, bet gan visa organisma alkatības kaušanas process. I.I.
Pretēji I. I. Mečņikova teorijai uz XX gadsimta vālītes. bija vairāk patologu, smirdoņu lauskas izraisīja fagocitozi iekaisušajās bedrēs, tobto. kaites leikocīti (gnіy) tika ārstēti ar slimību izraisošām, nevis slimīgām šūnām.
Turklāt diakoni zināja, ka fagocīti - baktēriju nesēji organismā, ir dzīvotspējīgi infekciju izplatīšanā. I.I.Mečņikova alu idejas piecēlās; mācības, aicinot zahisnі kitini, scho dіyut šādu rangu, "aprij kitiny". Daudzi jaunie franču kolēģi pavairo vikoristaty valrieksti no saknes ar tādu pašu nozīmi. I.I.Mechnikov pieņēma šo iespēju un ieviesa terminu "fagocīts".
Robotika un Mečņikova teorija tika lieliski garantēta L.
Pastērs un lūdza Ilju Illiču pratsyuvati no viņa institūta no Parīzes.
Mechnikov atklāj trīs svarīgas fagocītu spējas:
Aizsargā un attīra spēkus pret toksīniem, produktiem pret audiem, pret infekcijām;
Yaka pārstāv antigēnu funkciju uz šūnu membrānas;
Sekrēcijas spēks, kas ļauj redzēt enzīmu sekrēciju citās bioloģiskajās valodās.
Spiralizējoties uz qi trīs fagocītu dominanci, fagocitozi var raksturot kā trīs posmus:
Ķīmijtakss;
saķere;
endocitoze;
Klitīnās tiek novērots noliktavas fagocitozes opsonizācijas process.
Opsonīni tiek fiksēti uz detaļām un є ar veiksmīgu lanku no fagocītiskajām šūnām. Galvas opsonīni ir komplementu un imūnglobulīnu noliktavas. Tse nada klitiny augsta jutība pret fagocītiem un spriyaє їх iznischennu.
Endocitoze ar fagocītu vakuolu - fagosomu - pieņemšanu. Makrofāgu granulas un azurofilās un specifiskās neitrofilu granulas pārvietojas uz fagosomu un seko tai, redzot savu vietu fagosomu audos.
Poglinannya ir sarežģīts iekšējs šūnu process, kas veicina ATP ģenerēšanas mehānismus, specifisku glikolīzi un oksīdu fosforilēšanos makrofāgos.
Neitrofiliem ir brētliņu mikrobicīdu darbības režīms.
Kisnezalezhny pristrіy polygaє pie zbіlshennі vіlshennya skābs un glikozes іz sinhronā vignanny bioloģiski aktīvs nestabils rezultāti ieviešanu skābs. Kisnevnezalezhny savienojuma mehānisms ar galveno katjonu proteīnu un lizosomu enzīmu klātbūtni, kas degranulācijas laikā šūpojas ap fagosomu.

Foto: Nathan Reading
Erliha imunitātes teorija- viena no pirmajām antiantivielu ražošanas teorijām, kuras pamatā ir fakts, ka klitīniem ir antigēnam specifiski receptori, kas vibrē kā antivielas pret antigēnu.
Paula Erliha rakstā autors asins pretmikrobu runu nosauca ar terminu "antiviela", ka baktērijas tajā laikā sauca ar terminu "korper" - mikroskopisks ķermenis.
Ale P. Erlich "redzēja" vairāk teorētisko ieskatu. Neatkarīgi no tiem, kuri tajā pašā laikā apzinājās, ka radījuma asinīs, kas nesaskaras ar konkrētu mikrobu, vai cilvēkiem neuzrāda antivielas pret konkrēto mikrobu, P. Erlihs it kā pēc kārtas ieraudzīja. ka jau pirms saskarsmes ar konkrēto mikrobu organismā redzēju, kā vin tos sauca par "bičny lanceriem".
Kā mēs zinām tagad, bet tāpat, Erliha "bichni lansyugs" ir ziņots, ka ir izstrādājuši limfocītu receptorus antigēniem. Pіznіshe tsey sposіb dumok P.
Erlihs "zastosuvavs" uz farmakoloģiju: savā vīnu ķīmijterapijas teorijā viņš pārnesa receptoru pārnesi uz ārstnieciskām runām organismā.
1908. gadā lpp. P. Erliham tika piešķirta Nobela prēmija par imunitātes humorālo teoriju.
Bezridkas imunitātes teorija- Teorija, kas izskaidro ķermeņa inficēšanos zemu infekciozu slimību veidā, ko izraisa klitīna specifiskā makulas rezistence pret slimiem.
Pamācoša imunitātes teorija- Nosauktais anti-sējums ir nosaukts, zgishi, loma izhmuni vidpovіdi ir antigēns, jaku matricas liktenis īpaši antidetektora formulēšanas laikā, zlomīni slēpj zmіnieglobiye,
Mečņikova imunitātes teorija ir teorija, aiz kuras antibakteriālās imunitātes vissvarīgākā loma slēpjas fagocitozē.
Spopčatka I.I. Mečņikovs kā zoologs eksperimentāli redzēja Melnās jūras bezmugurkaula jūras faunu Odesā un pievērsa uzmanību tiem, kas dziedē šūnas (celolocīti) un radības, kas aptraipa trešās puses vielas (cietās daļiņas un baktērijas), kas iekļuva iekšējā vidū.
Potim vіn pobachiv analoģija mіzh tsim yavishche un poglenannymi blіmi kіtiny asins skriemeļu radības mikrobu ķermeņiem. Šiem procesiem līdz I. I. Mečņikovam sekoja cita mikroskopija. Ale tіlki I.I.Mečņikovs redzēja, ka šī parādība nav viena sālījuma ēšanas process, bet gan visa organisma alkatības kaušanas process.
I.I. Pretēji I. I. Mečņikova teorijai uz XX gadsimta vālītes. bija vairāk patologu, smirdoņu lauskas izraisīja fagocitozi iekaisušajās bedrēs, tobto. kaites leikocīti (gnіy) tika ārstēti ar slimību izraisošām, nevis slimīgām šūnām.
Turklāt diakoni zināja, ka fagocīti - baktēriju nesēji organismā, ir dzīvotspējīgi infekciju izplatīšanā. I.I.Mečņikova alu idejas piecēlās; mācības aicinot zahisnі kitiny, scho deyut šādu rangu "aprij kitiny". Daudzi jaunie franču kolēģi mudināja uzvarošās valriekstu saknes ar tādu pašu nozīmi.
I. I. Mechnikov pieņēma šo iespēju un ieviesa terminu "fagocīts". Šos robotus un Mečņikova teoriju lieliski pagodināja L. Pastērs, un viņš lūdza Ilju Illihu uzņemt viņa institūtā netālu no Parīzes.
Imunitātes klonālās atlases teorija.
Bērneta teorija – teorija, kādēļ organismā tiek vainoti klitīna kloni, imūnkompetenti un dažādi antigēni; antigēns ir vibrācijas kontaktā ar dzīvotspējīgu klonu, stimulējot ar to antivielu veidošanos.
Šo teoriju sadalīja Frenks Bērnets (1899-1985), lai izskaidrotu funkciju imūnsistēma.
Imunna vіdpovid var liecināt par lielu antigēnu skaitu.
Cilvēka ķermenis ir vainojams simtiem tūkstošu antivielu molekulu sintezēšanā no dažādiem reģioniem, kuras tiek atpazītas.
Klonālās atlases teorija apstiprina:
1. Antivielas un limfocīti ar nepieciešamo specifiskumu organismā ir jau pirms pirmā kontakta ar antigēnu.
2. Limfocītiem, kas piedalās imūnsistēmā, uz membrānas virsmas ir antigēnam specifiski receptori.
B-limfocītu ar receptoriem gadījumā ir tādas pašas specifikas molekulas, kuras antivielas, tāpat kā limfocīti, dabiski ražo un izdala.
Ādas limfocītu virsmas receptoriem ir tikai viena specifika.
4. Limfocīti, sensibilizēti ar antigēnu, iziet cauri proliferācijas stadijām un veido lielisku plazmas šūnu klonu.
Plazmas šūnas sintezē antivielas ar mazāku specifiskumu, un tam tika ieprogrammēts limfocītu priekšējais virziens.
Vairošanās signāli ir citokīni, kurus redz citi klitīni. Limfocīti bieži var redzēt arī citokīnus.
Pateicoties šim klonālās atlases mehānismam, antivielas var uzkrāties augstā koncentrācijā, lai efektīvi cīnītos ar infekciju.
Līdzīgu mehānismu izmanto antigēnu specifisko T-limfocītu atlasei.
Proliferējošam klonam ir vajadzīga stunda, lai izveidotu pietiekamu skaitu šūnu.
Kuras ass skaņas pāriet dažas dienas pēc saskares ar antigēnu, pirmo reizi asinīs tiek konstatētas antivielas. Oskіlki tsі antitіla pazuda pēc antigēnu infūzijas, mēs runājam par nabutu imunnu vіdpovіd.
Gruntēto limfocītu populācijas pieauguma intensitāte palielinās, vēl svarīgāk, ar klitīna palielināšanos, veidojot antigēnu stimulu.
Šajā gadījumā ir vainojama mehānismu kombinācija, kas ietver antigēna saglabāšanu, limfocītu populācijas izveidošanu un pakāpenisku klitīna klonu skaita samazināšanu, kas nodrošina imūnsistēmas veselīgu atmiņu. (undbany).
Viens no efektīvākajiem kontroles mehānismiem ir tas, ka reakcijas produkts vienlaikus kalpo kā inhibitors. Tāda paša veida negatīva seruma reakcija var rasties, apstiprinot antivielas.
Ērliha imunitātes teorija ir viena no pirmajām antivielu veidošanās teorijām, kuras pamatā ir fakts, ka šūnām ir antigēnam specifiski receptori, kas vibrē kā antivielas pret antigēnu.
Paula Erliha rakstā autors asins pretmikrobu runu nosauca ar terminu "antiviela", baktēriju lauskas tolaik tika sauktas ar terminu "korper" - mikroskopiskās antivielas.
Ale P. Erlich "redzēja" vairāk teorētisko ieskatu. Neatkarīgi no tiem, par kuriem tika liecināti šīs stundas fakti, ka radījuma asinīs, kas nesaskaras ar konkrētu mikrobu, vai cilvēkiem nav antivielas pret konkrēto mikrobu, P. Erlihs it kā pēc kārtas, kam redzēju, ka pat pirms kontakta ar konkrētu mikrobu organismā, es skatījos uz priekšu, saucot tos par "bіchniy lancers". Kā mēs tagad zinām, tas ir tāpat, un Erlihs - tse, kā ziņots, mūsu stundā limfocītu receptori ir pret antigēniem. Tas pats domāšanas veids P.
Erlihs "zastosuvavs" uz farmakoloģiju: savā vīnu ķīmijterapijas teorijā viņš pārnesa receptoru pārnesi uz ārstnieciskām runām organismā. 1908. gadā lpp. P. Erliham tika piešķirta Nobela prēmija par imunitātes humorālo teoriju.
Pasters propagandēja izsmeltas varas teoriju; acīmredzot "nepieņemamības" teorijai ir nometne, kurā cilvēka ķermenis (tāpat kā dzīvības nesējs) neatbalsta mikrobu attīstību.
Prote autors ātri saprot, ka šī teorija nevar izskaidrot zemo brīdinājumu. Zokrema, Pastērs rādīja kā inficēt vistu ar sibīriju un apgriezt kājas aukstā ūdenī, tad viņai rodas slimība (lielākajiem prātiem vistas Sibīrijai ir nepieņemamas). Parādības attīstību izraisīja ķermeņa temperatūras pazemināšanās par 1-2 ° C, tāpēc nevarēja būt nekāda iemesla dzīvības barotnes izsīkšanai organismā.
Robota pagriešana (1. nodaļa)
285178510795001. Zīmējums pēc zinātniskās metodes, lai ilustrētu attēla sižetu
holandiešu mākslinieka J. Stīna "Pulss", kas sarakstīts 17. gadsimta vidū? 1) modelēšana 2) vimir
3) eksperiments 4) piesardzība
2. Kāds ir veids, kā vikoristovuetsya zem mikroskopa iet amēbu mežonis?
1) vimir 2) modelēšana
3) apsardze 4) apsardze
3.
Kā nosaukt zinātni, kas izstrādā likumus vēsturiskā attīstība
organiskā pasaule?
1) anatomija 2) evolūcija 3) ģenētika 4) ekoloģija
4.
Kurš tiek uzskatīts par imunitātes klitīna teorijas radītāju?
1) Č.Dārvins 2) I.P. Pavlova 3)L. Pasters 4) I.I. Mečņikovs
5. Dziedošajā zinātnes galerijā dziļāko zināšanu sistēma ir
1) fakts 2) eksperiments 3) teorija 4) hipotēze
6. Formulējiet hipotēzi - vidējo
1) apkopot faktus 2) parādīt attaisnojumu
3) apstiprināt datu ņemšanas objektivitāti 4) veikt eksperimentu
7.
Citoloģijas zinātne ir attīstījusi savu attīstību
1) evolūcijas teorija 2) klitīna teorija
3) refleksu teorija 4) gēnu teorija
8. Sistemātika ir zinātne, kas
1) organismu funkcijas dabā 2) strīdīgās saites starp organismiem
3) organismu dzīvesveids 4) organismu dzīve
9.
Samaziniet ķermeņa komplekta zīmi
1) I.P. Pavlovs 2) I.I. Mehņikovs 3)G. Mendelis 4) Č. Darvins
10. Kas ir zinātne par fotosintēzes procesu?
1) ģenētika 2) fizioloģija 3) ekoloģija 4) sistemātika
11. Konstatēts radību sezonālās molēšanas fakts
1) mikrokopēšanas metode 2) apsardzes metode
3) eksperimentālā metode 4) hibrīda metode
12. Precīzi iestatiet šķelto vplyu, pievienojot roslīna augšanu, varat
metodi
1) eksperiments 2) modrība 3) modelēšana 4) analīze
13.
Recesijas zīmju pārraides modeļi
1) ģenētika 2) antropoloģija 3) ekoloģija 4) molekulārā bioloģija
14. Kā zinātne izrok mirušo organismu atliekas?
1) paleontoloģija 2) ģenētika 3) embrioloģija 4) sistemātika
15.
Shēmu, atzveltnes krēslu, dabas līdzīgu objektu veidošana
metožu grupai
1) modelēšana 2) simulācija 3) piesardzība 4) eksperimentāls16.
310578513716000Kāda dzīves organizēšanas diena ir attēlota šajā fotogrāfijā?
1) molekulārā ģenētiskā
2) organoīds-klitīns
3) biogeocenotisks
4) populācija-sugas
17. 310642012255500 Kāda dzīves organizācija ir projicēta šim mazajam?
1) molekulārā ģenētiskā
2) organoīds-klitīns
3) organisms
4) biogeocenotisks
18. Kāda dzīvo būtņu organizācija ir galvenais citoloģijas attīstības objekts?
1) bioģeocenotiskā 2) populācija-suga
3) clinny 4) biosfērisks
19.
Kā eksperimenta metode atšķiras ar piesardzības metodi?
2) vīni tiek veikti speciāli radītos un kontrolētos prātos
3) vairāk braucienu vienā pavadītajā stundā
4) joga
20. Kā modelēšanas metode atšķiras ar piesardzības metodi?
1) no sertifikātiem tiek ņemts jogas vadīšanas process zinātniskiem faktiem
2) joga, lai vadītu kvalifikācijas apmācību
3) laimē vairāk trivaļu pa stundām
4) netiek kopēts pats objekts, piemēram, kopija
21.
Yaka zinātne vvchaє vnutrishnyovidov vzaєmini organіzmіv:
1) sistemātika 2) ekoloģija 3) selekcija 4) morfoloģija
22. Kādā līmenī ir dzīvo būtņu organizācija dabā
runu loks?
1) klitīns 2) organisms
3) populācija-sugas 4) biosfēras
23.
Kādā dzīves organizācijas līmenī notiek cīņa par pamatu starp iedzīvotājiem?
1) sugas 2) organisms 3) biocenotisks 4) biosfērisks
24. Dzīviem dabas objektiem, nedzīvu ķermeņu skatam tas raksturīgs
1) mainot vagonu 2) pārvietojoties tuvu atklātai telpai
3) elpošana 4) runāšana pie ūdens
25.Dūmu lapsas zīlīšu viendabīga grupa - vienlīdzīgas organizācijas dibens
dzīvs
1) organisms 2) biosfēra
3) bioģeonotiskā 4) populācija-sugas
26.
Cik lielā mērā dzīves organizācija izseko pēctecībai
planētas ekosistēmas?
1) specifisks; 2) biosfēra; 3) iedzīvotāju skaits;
27.
Červonas stallis, kas aizņem dziedāšanas laukumu, ir dzīvās dabas organizācijas plosīts.
1) organisms 2) biocenotisks
3) biosfēras 4) populācijas-sugas
28. Olbaltumvielu biosintēzes process notiek tikai uz vienādiem noteikumiem:
1) organisms 2) molekulārais
3) biosfēras 4) populācijas-sugas
29.
Visvienlīdzīgākā dzīves organizācija;
1) organismi 2) molekulārie
3) biosfēras; 4) biogeocenotisks.
30. Roslīnu pamatšķirņu un šķirņu uzlabošana
nodarbojoties ar zinātni
1) ekoloģija 2) molekulārā bioloģija 3) selekcija 4) ģenētika
31. Viena no dzīvā un nedzīvā klātbūtnes pazīmēm ir ēkas integritāte pret
1) mainīt rozmiriv 2) pašrade
3) sagraut; 4) bezbiržas pieaugums
32.
Visi dzīvie organismi apvienojas:
1) clitina budova 2) ēka pirms fotosintēzes
3) kodola parādīšanās klitīnā 4) ēka sagraut
33. Visiem dzīviem organismiem, veidojot:
1) pārvietoties 2) mainīt vārdus
3) olbaltumvielu, tauku, ogļhidrātu ēšana 4) neraudzēta augšana
34. Izplatījis dzīvo organismu klasifikācijas sistēmu un sakārtojis sugu bināro nomenklatūru:
1) G. Mendels 2) K. Linnijs 3) J. B. Lamarks 4) I. Mečņikovs
Mečņikova imunitātes teorija- teorija, kurai antibakteriālās imunitātes loma ir fagocitozē.
Spopčatka I.I. Mečņikovs kā zoologs eksperimentāli redzēja Melnās jūras bezmugurkaula jūras faunu Odesā un pievērsa uzmanību tiem, kas dziedē šūnas (celolocīti) un radības, kas aptraipa trešās puses vielas (cietās daļiņas un baktērijas), kas iekļuva iekšējā vidū. Potim vіn pobachiv analoģija mіzh tsim yavishche un poglenannymi blіmi kіtiny asins skriemeļu radības mikrobu ķermeņiem. Šiem procesiem līdz I. I. Mečņikovam sekoja cita mikroskopija. Ale tіlki I.I.Mečņikovs redzēja, ka šī parādība nav viena sālījuma ēšanas process, bet gan visa organisma alkatības kaušanas process. I.I. Pretēji I. I. Mečņikova teorijai uz XX gadsimta vālītes. bija vairāk patologu, smirdoņu lauskas izraisīja fagocitozi iekaisušajās bedrēs, tobto. kaites leikocīti (gnіy) tika ārstēti ar slimību izraisošām, nevis slimīgām šūnām. Turklāt diakoni zināja, ka fagocīti - baktēriju nesēji organismā, ir dzīvotspējīgi infekciju izplatīšanā. I.I.Mečņikova alu idejas piecēlās; mācības, kas činni sauc par tādu rangu zahisni klitīni "aprij klitīnus". Daudzi jaunie franču kolēģi pavairo vikoristaty valrieksti no saknes ar tādu pašu nozīmi. I.I. Mečņikovs, pieņemot šo iespēju, un parādījis terminu "Fagocīts".Šos robotus un Mečņikova teoriju lieliski pagodināja L. Pastērs, un viņš lūdza, lai Illija Illiča tiktu sertificēta viņa institūtā Parīzē.
Erliha imunitātes teorija- viena no pirmajām antiantivielu ražošanas teorijām, kuras pamatā ir fakts, ka klitīniem ir antigēnam specifiski receptori, kas vibrē kā antivielas pret antigēnu.
Paula Erliha rakstā autors asins pretmikrobu runu nosauca ar terminu "antiviela", ka baktērijas tajā laikā sauca ar terminu "korper" - mikroskopisks ķermenis. Ale P. Erlich "redzēja" dziļu teorētisko ieskatu. Neatkarīgi no tiem, par kuriem liecināja šīs stundas fakti, ka radījumu vai cilvēku asinīs, it kā tie nav nonākuši saskarē ar konkrētu mikrobu, antivielas pret šo mikrobu netiek konstatētas, P. Erlihs it kā uzzināja. ka pat pirms saskares ar konkrētu mikrobu organismā. jau є antityla vigliādos, kurus viņš nosauca par "bičnija lanceriem". Kā mēs tagad zinām, tas ir tāpat, un Erliha "bichni lansyugs" - tas ir ziņojums mūsu stundā limfocītu antigēnu receptori. P.Ērliha jaunais domāšanas veids "zastosuv" uz farmakoloģiju: savā vīnu ķīmijterapijas teorijā viņš pārnesis receptoru pārnesi organismā uz ārstnieciskām runām. 1908. gadā lpp. P. Erliham tika piešķirta Nobela prēmija par humorālā imunitātes teorija.
Ir arī dažas citas teorijas.
Bezridkas imunitātes teorija- teorija, kas izskaidro ķermeņa inficēšanos zemas infekcijas slimību veidā, ko izraisa klitīna specifiskā makulas rezistence pret slimībām.
Pamācoša imunitātes teorija- Nosauktais anti-sējums ir nosaukts, zgishi, loma izhmuni vidpovіdi ir antigēns, jaku matricas liktenis īpaši antidetektora formulēšanas laikā, zlomīni slēpj zmіnieglobiye,
№69 Pretvīrusu, antibakteriālas, pretsēnīšu, pretaudzēju, transplantācijas imunitātes pazīmes.
Pretvīrusu imunitāte. Pretvīrusu imunitātes pamats ir šūnu imunitāte. Ar vīrusu inficētas šūnas, kas satur citotoksiskus limfocītus, kā arī NK šūnas un fagocītus, kas mijiedarbojas ar vīrusam specifiskiem proteīniem inficētajās šūnās piesaistīto antivielu Fc fragmentiem. Veselības pretvīrusu antivielas mazāk neitralizē pozaklitinno roztashovanі vīrusus, kā nespecifiskās imunitātes faktoru - syrovatkovі anti-rusnі ingіbіtori. Šādus vīrusus, kas noslīpēti un bloķēti ar olbaltumvielām ķermenim, tiek iztīrīti ar fagocītiem, vai arī tie tiek izņemti no sekcijas, tad tas. (Tātad nosaukumi ir "redzama imunitāte"). Interferoniem piemīt pretvīrusu rezistence, izraisot enzīmu sintēzi klitīnās, kas kavē nukleīnskābju un vīrusu proteīnu veidošanos. Krym tsgogo, іninterferoni remonts imunomodulyuyuyuyu dіyu, polyuyut in klitiny ekspresіyu antigenov galvas kompleksā histosummіnostі (MHC). Gļotādu pretvīrusu aizsardzība ar sekrēcijas IgA, kas kopā ar vīrusiem maina to saķeri ar epitēliocītiem.
Antibakteriālā imunitāte vērsta pret baktērijām un pret toksīniem (antitoksiska imunitāte). Baktērijas un to toksīnus neitralizē antibakteriālas un antitoksiskas antivielas. Baktēriju (antigēna)-antivielu kompleksi aktivizē komplementu, kura sastāvdaļas ir pievienotas antivielas Fc fragmentam, un pēc tam izveido membrānas uzbrukuma kompleksu, kas iznīcina gramnegatīvo baktēriju šūnu sienas ārējo membrānu. Baktēriju šūnu sieniņu peptidoglikānu iznīcina lizocīms. Antivielas un komplements (C3b) apņem baktērijas un “pielīmē” tās pie Fc- un C3b receptoriem fagocītos, spēlējot opsonīnu lomu kopā ar citiem proteīniem, kas uzlabo fagocitozi.
Galvenais antibakteriālās imunitātes mehānisms ir fagocitoze. Fagocīti tieši pāriet uz fagocitozes objektu, reaģējot uz ķīmijatraktantiem: mikrobu runu, aktivētā komplementa komponentiem (C5a, C3a) un citokīniem. Gļotādu antibakteriāla infekcija ar sekretoro IgA, kas kopā ar baktērijām maina to saķeri ar epitēliocītiem.
Pretsēnīšu imunitāte. Antivielas (IgM, IgG) tiek noteiktas zemā titrā mikozes gadījumā. Pretsēnīšu imunitātes pamats ir šūnu imunitāte. Audos notiek fagocitoze, attīstās epitēlija granulomatoza reakcija un dažreiz asinsvadu tromboze. Mikozes, īpaši oportūnistiskas, bieži attīstās pēc izmēģinājuma antibakteriālās terapijas un imūndeficīta gadījumā. Smaku pavada uzlabotā tipa paaugstinātas jutības paaugstināšanās. Iespējama alerģisku slimību attīstība pēc elpceļu sensibilizācijas ar Aspergillus, Penicillium, Mucor, Fusarium u.c. prātu patogēno sēņu nojumes fragmentiem.
Pretpietūkuma imunitāte pamats Th1 depozīta šūnu imūnās atbildes reakcijai, kas aktivizē citotoksiskos T-limfocītus, makrofāgus un NK-šūnas. Humorālās (antilas) imūnās atbildes loma ir maza, antivielu lauskas, kas saistītas ar apaļo šūnu antigēnu determinantiem, pārbauda to citopatogēno imūno limfocītu veidus. Pūkaino antigēnu atpazīst antigēnu prezentējošie klitīni (dendrītiskie klitīni un makrofāgi), un bez starpnieka vai ar T-palīgu (Th1) tiek parādīts citotoksiskajiem T-limfocītiem, kas iznīcina kuplo klitīna mērķi.
Specifiskas pretvēža imunitātes imūnnovērošana normālai audu noliktavai tiek realizēta nespecifisku faktoru dēļ. Nespecifiski faktori, kas stimulē puffer šūnas: 1) NK-šūnas, mononukleāro šūnu sistēma, kuras pretvēža aktivitāti pastiprina interleikīna-2 (IL-2) un α-, β-interferonu pieplūdums; 2) LAK šūnas (mononukleārās šūnas un NK šūnas, ko aktivizē IL-2); 3) citokīni (α- un β-interferons, FNP-α un IL-2).
Transplantācijas imunitāte nosaukt imūnreakciju pret makroorganismu, kas vērsta pret pārstādītu uz jaunu svešu audu (transplantu). Zināšanas par transplantācijas imunitātes mehānismiem ir nepieciešams risinājums vienai no mūsdienu medicīnas svarīgākajām problēmām - prokuratūras orgānu un audu transplantācijai. Labs ziņojums, kas parāda, ka svešu orgānu un audu transplantācijas operācijas panākumi svarīgākajos gadījumos ir saistīti ar donora un recipienta audu imunoloģisko integritāti.
Imūnā reakcija uz svešām šūnām un audiem ir saistīta ar to, ka antigēni, kas organismam ir ģenētiski sveši, atrodas viņu noliktavā. Qi antigēni, kas atņēma transplantācijas nosaukumu vai antigēnus histosumijā, ir visvairāk pārstāvēti CPM klitīnā.
Tirgotāja reakcija netiek vainota donora un recipienta kopuma gadījumā par antigēniem histosumijā - tas var būt mazāks par identiskajiem dvīņiem. Reakcijas nopietnība uz invāziju ir bagāta ar to, ko apgulties svešuma stadijā, uz materiālu, kas tiek pārstādīts, un es kļūs par recipienta imūnreaktivitāti.
Saskaroties ar svešiem transplantācijas antigēniem, organisms reaģē ar šūnu un humorālās imunitātes faktoriem. Galvenais faktorsšūnu transplantācijas imunitāte є T-killer. Šūnas pēc sensibilizācijas ar donora antigēniem migrē uz transplantāta audiem un izraisa pretšūnu izraisītu citotoksicitāti uz tām.
Transplantācijas imunitātes veidošanā svarīgas var būt specifiskas antivielas, kas saistās ar svešiem antigēniem (hemaglutinīniem, hemolizīniem, leikotoksīniem, citotoksīniem). Tie izraisa transplantāta citolīzes starpniecību pret antivielām (komplementa mediācija un antivielu šūnu izraisīta citotoksicitāte).
Iespējama adaptīva transplantācijas imunitātes pārnešana papildus aktivizētiem limfocītiem vai specifiskam pretkairinājuma līdzeklim sensibilizētam neskarta makroorganisma indivīdam.
Šūnu un audu transplantācijas imūnsupresijas mehānismam var būt divas fāzes. Pirmajā fāzē, netālu no transplantāta, šis trauks ir pakļauts imūnkompetentu šūnu uzkrāšanai (limfātiskā infiltrācija), tostarp T-killers. Otrā fāzē tiek novērota šūnu transplantāta iznīcināšana ar T-killeriem, tiek aktivizēta makrofāgu lanka, dabiskais slepkava un specifiska antitiloģenēze. Vainot imūno iekaisumu, asinsvadu trombozi, transplantāta norīšanas traucējumus un nāvi. Izpostītos audus izmanto fagocīti.
Reakcijas procesā veidojas imūnās atmiņas T-B-šūnu klons. Atkārtota šo orgānu un audu transplantācijas pārbaude atklāj otru imūnreakciju, jo tā plūst rīstošāk un ātri beidzas ar transplantāta atgrūšanu.
No klīniskā viedokļa plaisa tiek uzskatīta par akūtu, pārkarsētu un pārmērīgi izstieptu transplantātu. Smaku rada reakcijas īstenošanas stunda un citi mehānismi.
Mečņikova imunitātes teorija - teorija, kurai antibakteriālās imunitātes loma ir fagocitozē.
Spopčatku I.I. Mečņikova jaku zoologs eksperimentāli vivchav jūras bezmugurkaula faunaMelnās jūras krastā Odesā un izrādot cieņu tiem, kuridilenі kіtini (coelomocytes) tsikh radījumi pūttrešās puses vielas (cietās daļiņas un baktērijas), kas ir iekļuvušas iekšējā kodolā. Potim vіn pobachiv analoģija mіzh tsim yavishche un poglenannymi blіmi kіtiny asins skriemeļu radības mikrobu ķermeņiem. Šiem procesiem līdz I. I. Mečņikovam sekoja cita mikroskopija. Ale tilkilīdz I.I.zhivlennya tsієї vientuļa klitīna un є zahisniy processuz visa organisma rēķina. I.I.plaisāšana iekaisusi kā zahisne, bet ne graujoša izpausme. Pretēji I.I.Mečņikova uz vālītes teorijai XX iekšā. bija vairāk patologu, smirdoņu lauskas izraisīja fagocitozi iekaisušajās bedrēs, tobto. kaites leikocīti (gnіy) tika ārstēti ar slimību izraisošām, nevis slimīgām šūnām. Turklāt nēyakі vvazhali, scho fagocīti - baktēriju nesēji organismā, vidpovidalnі infekciju izplatīšanai. Alus idejasI.I.Mečņikovs piecēlās kājās; nosaucot tādas lietasrangs zahisni klitīni "aprij klitīnus". Jogo moFranču kolēģu kori mudināja vikorēt griķusnav tādas pašas nozīmes saknes. I.I. Mečņikovs, pieņēmis misijuriant, i z'yavsya term "Fagocīts". Qi robotu teorijaMečņikovu pagodināja L.Pasters, un lūdzaIllu Illich pratsyuvati spēki savā institūtā netālu no Parīzes.
Erliha imunitātes teorija - Viena no pirmajām antiantivielu ražošanas teorijām, saskaņā ar kuru klitīniem ir antigēnu specifiski receptori, kas vibrē kā antivielas pret antigēnu.
Pie Pāvila Erliha statujasasins autora pretmikrobu runasaucot terminu "antiviela", lauskasbaktērijas tajā laikā sauca par terminu " korper "- mikro pichnі tіltsya. Ale P. Erlich "redzēja" dziļu teorētiskogodīgi. Nezināja par tiem, kuri patiesībā bija liecinieki tajā stundā, ka asinīs viņi nesaskārās ar viltuRadības ir noteikts mikrobs, bet cilvēki to neapzīmēantivielas pret konkrēto mikrobu, P. Erlihs pārliecinājās, ka pirms saskares ar konkrētu mikrobu organismāes jau є antivielas redzeslokā, ko vin sauca par "bichnymi".lancers". Kā mēs tagad zinām, tas ir tāpat, un "puseYerlikha's lancers - tse ziņojums vvcheni mūsu stundā recepte tori limfocīti antigēniem Pіznіshe tsey pats domāšanas veidsP. Erlihs "zastosuvavs" uz farmakoloģiju: savā ķīmijas teorijāvīnu terapijas, kas pārnesušas receptes pamattekstāārstnieciskām runām. 1908. gadā lpp. P. Erlihs tika apbalvotsNobela prēmija par humorālā imunitātes teorija.
Ir arī dažas citas teorijas .
Bezridkas imunitātes teorija - teorija, kas izskaidro ķermeņa inficēšanos zemas infekcijas slimību veidā, ko izraisa klitīna specifiskā makulas rezistence pret slimībām.
Instrukcija un imunitātes teorijā - Nosauktais anti-sējums ir nosaukts, zgishiy, loma izhmuni vidpovіdi ir antigēns, un anchstye matricas liktenis konfigurācijas faktora formulējumā, čūska, slēpjas ar čūsku.
Ir zināms, ka klitīniem ir specifiski antigēnu receptori, kas vibrē kā antivielas pret antigēnu.
Paula Erliha rakstā autors asins pretmikrobu runu nosauca ar terminu "antiviela", ka baktērijas tajā laikā sauca ar terminu "korper" - mikroskopisks ķermenis. Ale P. Erlich "redzēja" vairāk teorētisko ieskatu. Neatkarīgi no tiem, par kuriem liecināja šīs stundas fakti, ka radījumu vai cilvēku asinīs, it kā tie nav nonākuši saskarē ar konkrētu mikrobu, antivielas pret šo mikrobu netiek konstatētas, P. Erlihs it kā uzzināja. ka pat pirms saskares ar konkrētu mikrobu organismā. jau є antityla vigliādos, kurus viņš nosauca par "bičnija lanceriem". Kā mēs tagad zinām, tas ir tāpat, un Erliha "bichni lansyugs" - tas ir ziņojums mūsu stundā limfocītu antigēnu receptori. P. Erliha jaunais domāšanas veids "zastosuv" uz farmakoloģiju: savā vīnu ķīmijterapijas teorijā viņš pārnesa receptoru pārnesi organismā uz ārstnieciskām runām. 1908. gadā lpp. P. Erliham tika piešķirta Nobela prēmija par humorālā imunitātes teorija.
Ir arī dažas citas teorijas.
Bezridkas imunitātes teorija - teorija, kas izskaidro ķermeņa inficēšanos zemas infekcijas slimību veidā, ko izraisa klitīna specifiskā makulas rezistence pret slimībām.
Pamācoša imunitātes teorija - Nosauktais anti-sējums ir nosaukts, zgishi, loma izhmuni vidpovіdi ir antigēns, jaku matricas liktenis īpaši antidetektora formulēšanas laikā, zlomīni slēpj zmіnieglobiye,
28) Imunoloģiskā tolerance, mehānismi
Imunoloģiskā tolerance- Vіdsutnіst spetsifichnії produktīvs іmunnoї vіdpovіdі organisms uz antigēna vіz'yazku z nezdatnіstyu yogo razznіvannya. Runājot par imūnsupresiju, imunoloģiskā tolerance pārnes vālītes nereaģēšanu uz pirmo antigēnu.
Vlasne imunoloģiskās tolerances fenomens tika atklāts 1953. gadā. neatkarīgais zinātnieks M. Hašeks un angļu zinātnieku grupa šajā jomā kopā ar P. Medavaru. Gashok gadījumos ar vistu embrijiem un Medavar jaundzimušajām pelēm parādīja, ka organisms kļūst nejutīgs pret antigēnu, kad to ievada embrionālajā vai agrīnā pēcdzemdību periodā.
Imunoloģiskā tolerance izsauc antigēnus, jaki atņēma nosaukumu tolerogēni. Tās var būt praktiski visas runas, polisaharīdi var būt vistolerogēnākie.
Imunoloģiskā tolerance ir iedzimta un pildīta. dibens iedzimtā toleranceє vіdsutnіst imūnsistēmas reakcija uz organisma antigēniem. Nabutu tolerance iespējams izveidot veidu, kā ievadīt antigēnu embrija periodā vai pirmajās dienās pēc indivīda piedzimšanas.
Imunoloģiskā tolerance ir atkarīga no specifikas - tā tiek iztaisnota, lai stingri dziedātu antigēnus. Aiz platuma pakāpes ir daudzvērtīga un dalīta tolerance. Polivalenta imunoloģiskā tolerance vinikaє vienu stundu uz visiem antigēnu determinantiem, kas nonāk konkrēta antigēna noliktavā. Priekš sadalīt, vai monovalence, tolerance tipiski viborcha nespriyyatlivost okremih antigēnu noteicošie faktori.
Imunoloģiskās tolerances izpausmes stadija pašlaik tiek nogulsnēta maza makroorganisma un tolerogēna jaudas klātbūtnē. Tātad, parādoties tolerancei, tas tiek ievadīts ķermeņa imūnreaktivitātes stāvoklī. Imunoloģiskā tolerance ir vieglāk inducējama embrionālajā periodā un pirmajās dienās pēc piedzimšanas, tā ir visizteiktākā radījumiem ar samazinātu imūnreaktivitāti un ar vienādu genotipu.
Imunoloģiskās tolerances indukcijas panākumi ir atkarīgi no antigēna svešuma pakāpes organismam, ņemot vērā tā raksturu, zāļu devu un antigēna injekcijas uz ķermeņa trivalitāti. Vislielākā tolerogenitāte var būt vismazāk svešķermeņu antigēnu iedarbības ziņā, kam var būt maza molekulmasa un augsta viendabība. Vieglāk ir veidot toleranci pret aizkrūts dziedzera neatkarīgiem antigēniem, piemēram, baktēriju polisaharīdiem.
Atšķiriet lielas un mazas devas toleranci. Augstas devas tolerance pieprasot lielu skaitu augsti koncentrētu antigēnu. Kādos apstākļos pastāv tieša maldība starp runas devu un vibrācijas efektu. zemas devas tolerance, no otras puses, tas izsauc pat nelielu daudzumu ļoti viendabīga molekulārā antigēna. Devas iedarbība dažādos gadījumos var būt pretēja
Eksperimentā tolerance veidojas pēc dažām dienām, un dažreiz pēc tolerogēna ievadīšanas, kā likums, tā izpaužas ilgu laiku, vīnu doki cirkulē organismā. Efekts ir vājāks, vai arī tas ir pievienots tolerogēnam no ķermeņa. Skaņa imunoloģiskā tolerance nav triviāla stunda - mazāk nekā dažas dienas. Ilgstošai zāļu lietošanai ir nepieciešamas atkārtotas injekcijas.
Tolerances mehānismi ir dažādi un paliek neatšifrēti. Acīmredzot par pamatu vajadzētu būt normāliem imūnsistēmas regulēšanas procesiem. Imunoloģiskās tolerances attīstībai ir trīs galvenie iemesli: limfocītu antigēnu specifisko klonu izvadīšana no organisma; imūnkompetentu šūnu bioloģiskās aktivitātes bloķēšana; antigēnu neitralizācija ar antivielām.
Eliminācijas vai dzēšana, kā likums, ir autoreaktīvo T-limfocītu kloni to ontogēna agrīnā stadijā.
per. Antigēnam specifiska receptora aktivizēšana TCR nenobriedušu limfocītu indukcija jaunā apoptozē. Šī parādība, kas nodrošina organisma nereaktivitāti pret autoantigēniem, izlaižot nosaukumu centrālā tolerance. Vietējā tolerance pret barjeras antigēniem, audus aizsargā dabiskās šūnas, kas sensibilizē pret šiem T-limfocītu antigēniem.
Imūnkompetento šūnu bioloģiskās aktivitātes blokādes galvenā loma pieder imūncitokīniem. Vplivayuchi uz vіdpovіdnі receptoriem, smaka var izraisīt vairākas negatīvas sekas. Piemēram, T- un B-limfocītu proliferāciju aktīvi cinko β-TGF. T0-helpera diferenciāciju T 1 var bloķēt ar IL-4, 13 un T 2 palīga - γ-IFN palīdzību. Makrofāgu bioloģisko aktivitāti kavē T2 palīgprodukti (IL-4, 10, 13, β-TGF un citi).
Biosintēze B-limfocītos un tās pārvēršana plazmacītos ir saistīta ar brīvi cirkulējošu IgG. Antigēnu molekulu inaktivācija ar antivielām novērš to saistīšanos ar imūnkompetentu šūnu receptoriem - tiek izvadīts specifiskais aktivējošais faktors. Iespējams, adaptīva neskartu radījumu imunoloģiskās tolerances pārnešana, ieviešot imūnkompetentas šūnas, kas ņemtas no donora.
Imunoloģiskās tolerances fenomenam var būt liela praktiska nozīme. Vіn vikoristovuєtsya par vyrіshennya bagatioh svarīgas medicīnas problēmas, piemēram, orgānu un audu transplantācija, autoimūnu reakciju nožņaugšana, alerģiju un citu patoloģisku stāvokļu ārstēšana, kas saistīti ar imūnsistēmas agresīvu uzvedību.
Toleranci var apvienot. Šim nolūkam ir nepieciešams aktivizēt imūnsistēmu ar adjuvantiem, interleikīniem vai pārslēgt tiešo reakciju uz imunizāciju, modificējot antigēnus. Otrs veids ir tolerogēna izvadīšana no organisma, injicējot specifiskas antivielas vai imūnsorbcijas veidā.
29) Klitīna imūnreakcija (citotoksicitāte un aizdegšanās imūnreakcija, citokīnu, T-helperu un makrofāgu nozīme)
Klinta imunitāte- tas ir imūnās atbildes veids, kurā nepiedalās ne antivielas, ne komplementa sistēma. Šūnu imunitātes procesā tiek aktivizēti makrofāgi, dabiskās šūnas, antigēnu specifiskie citotoksiskie T-limfocīti, un antigēnā ir redzami citokīni.
Imūnsistēma vēsturiski ir sadalīta divās daļās - humorālās imunitātes sistēmā un kliniskās imunitātes sistēmā. Humorālās imunitātes gadījumā tiek likvidētas plazmā esošo molekulu funkcijas, bet ne šūnu elementi. Tāpat citā klitīna imūnsistēmā funkcija ir saistīta ar pašas imūnsistēmas klitīnu. CD4 diferenciācijas klastera vai T-palīgu šūnu limfocīti aizsargā pret dažādiem patogēniem.
Klīniskās imunitātes sistēma pārvar savas funkcijas šādos veidos:
antigēnu specifisko citotoksisko T-limfocītu aktivācijas veids, kas var izraisīt somatisko šūnu apoptozi, uz virsmas demonstrējot svešu antigēnu epitopus, piemēram, ar vīrusiem inficētas šūnas, kas atriebj baktērijas un šūnas, kurām ir apjomīgi antigēni;
makrofāgu un dabisko šūnu aktivācijas veids, kā iekšējo šūnu patogēnu iznīcināšanas veids;
veids, kā stimulēt citokīnu sekrēciju, ko ievadīt citās imūnsistēmas šūnās, kas piedalās adaptīvajā imūnreakcijā un iedzimtajā imūnreakcijā.
Citotoksiskie CD8+ CTL var nogalināt ar vīrusu inficētas šūnas, lai ar tām nonāktu tiešā saskarē. Saskares vietā ar CTL poras veidojošie proteīni, perforīni, iekļūst klitīna mērķa membrānā, līdzīgi kā membrānas mikrokanāli, kuru fermenti, fragmenti, iekļūst klitīna mērķī, izraisot klitīna kodola iznīcināšanu un nāvi.
30) Humorālā imūnreakcija (citokīnu nozīme, Th-2 limfocīti un B-limfocīti).
31) Antithila. Imūnglobulīnu klase, struktūra un funkcija.
Viena no senākajām imūninfekcijas filoģenētiskajām formām ir antivielu – proteīnu, kas specifiski reaģē ar antigēniem – biosintēze. Antivielas ir svarīgas proteīnu globulīna frakcijai asins plazmā, dažos gadījumos tās samazinās par 15-25% no olbaltumvielu satura, kas kļūst aptuveni 10-20 g / l. Tai antitilai viņi atņēma vārdu imūnglobulīni, tie їх ir apzīmēti ar simbolu Ig. Tās pašas antivielas - tie paši asins plazmas γ-globulīni, kas specifiski saistās ar antigēnu un piedalās dažādās imūnreakcijās.
Antivielas sintezē B-limfocīti un to pielikumi - plazmas šūnas - cirkulējošā formā, un receptoru molekulu veidā uz imūnkompetentām šūnām. Cirkulējošās antivielas ir iedalītas syrovatkovy un sekrēcijas.
Imūnglobulīni ir asins seruma proteīni. Smaku no specifiska antigēna izdala plazmas klitīni.
Papuve pēc molekulārās un svarīgās lancetes pazīmēm, kā arī izotipisko vai grupu antigēnu determinantu izpausmēm izšķir 5 Ig klases jeb izotipus (11.2. att.). Molekulas, kas atriebjas α tipam, tiek audzinātas līdz izotipam jeb A klasei (saīsināti IgA), δ tipam - IgD, ε tipam - IgE, γ tipam - IgG un μ tipam - IgM. Ir arī Ig apakšklase.
Ādas Ig izotipu raksturo specifiskas pazīmes. Zokrema, Ig D, E un G var būt monomēri, IgM praktiski ir pentamērs, bet IgA molekula var būt mono-, di-trimērs. Lielākais raksturīgie rīsi tabulā inducēti dažādi Ig izotipi. 11.1.
Imūnglobulīnu klase G lai kļūtu par galveno asinsrites sistēmas Ig masu, 70-80% no visa cirkulējošā Ig nokrīt uz її daļu, 50% nogulsnējot audos mājās. Vidējais IgG daudzums veselu nobriedušu cilvēku asinīs ir 12 g/l, kas sasniedz 7-10 gadu vecumu. IgG sabrukšanas periods ir 21 diena.
IgG ir monomērs, kas var saistīt 2 antigēnu saistošos centrus, kas var saistīt 2 molekulas ar antigēnu pospoli. Molekulmasa ir tuvu 160 kD, sedimentācijas konstante ir 7S. Sintezē nobrieduši B-limfocīti (γ) un plazmas šūnas. Labs ir norādīts asins serozā primārās imūnās atbildes pīķa un sekundārās imūnās atbildes gadījumā. Maє augsts afinitāte.
Atsevišķi apakštipi G1-G4. IgG1 un G3 saistās ar komplementu, un G3 ir aktīvāks. IgG4 ir līdzīgs IgE, var būt citofīls (tropisks vai sporīds, lai izlīdzinātu šūnas un bazofīlus) un piedalīties I tipa alerģiskās reakcijas attīstībā.
Pirmajos 3-4 mēnešos pēc dzemdībām ir viegli iziet cauri placentas barjerai un nodrošināt humorālo imunitāti jaundzimušajam, turklāt tie parādās pienā. IgG nodrošina neitralizāciju un antigēna marķēšanu, kas izraisa komplementa mediētu citolīzi un ADCC.
Imūnglobulīna klase M - Lielākā molekula no visiem Ig. Tse pentamērs, kuram var būt 10 antigēnu saistīšanas centri. Jogo molekulmasa ir tuvu 900 kDa, sedium konstante ir
mentalitāte 19S. Atsevišķi apakštipi M1 un M2. Smagas IgM molekulas lāpstiņas, pamatojoties uz citiem izotipiem, inducētas no 5 domēniem. Tā kā J-lance ir polimēra molekula, ir iespējams nomainīt. Periods napіvrozpadu 5 dienas.
5-10% no visa cirkulējošā Ig nokrīt uz šo daļu. Vidējais IgM daudzums vesela nobrieduša cilvēka asinīs ir tuvu 1 g/l. Tsgogo vienādi cilvēki syagaє jau 2-4-upes gadsimtā. IgM ir filoģenētiski senākais imūnglobulīns. Iekārtojies uz primārās imūnās atbildes vālītes.
Var būt ļoti atsaucīgs, visefektīvākais komplementa aktivators aiz klasiskā maršruta. Normālākas antivielas un izoaglutinīni nokļūst IgM. Chi neiziet cauri placentai. Augstu specifisku antivielu pret M izotipu titru noteikšana jaundzimušā asins serotipā liecina par pārmērīgu intrauterīnās infekcijas un placentas defektu. IgM nodrošina neitralizāciju un antigēna marķēšanu, izraisot komplementa mediētu citolīzi un ADCC. Є smaga infekcijas procesa marķieris.
Imūnglobulīna klase A іsnuє in syrovatkovіy un sekrēcijas formas. Gandrīz 60% no visa IgA ir atrodami gļotādu izdalījumos.
Sirovatkovijs IgA. Jogā bieži nokrīt gandrīz 10-15% no visa cirkulējošā Ig. Veselās asins šūnās veseliem pieaugušajiem ir aptuveni 2,5 g / l IgA, maksimums sasniedz 10. gs. Periods navіvrozpadu 6 dienas.
IgA ir monomērs, tam ir 2 antigēnu saistīšanas centri, molekulmasa aptuveni 170 kD un sedimentācijas konstante 7S. Atsevišķi apakštipi A1 un A2. Sintezē nobrieduši imūnie B-limfocīti (α) un plazmas šūnas. Labs ir norādīts asins serozā primārās imūnās atbildes pīķa un sekundārās imūnās atbildes gadījumā. Var būt augsta afinitāte. Chi neuzrāda papildinājumu. Chi neiziet cauri placentas barjerai. IgA nodrošina neitralizāciju un antigēna marķēšanu, kas izraisa ADCC sākumu.
sekretārs IgA (s IgA) ir polimēra formā šķietami dili trimerā (4 vai 6-valentā), kas satur 4 vai 6 paratopus un pat J- un S-peptīdus. Molekulmasa 350 kD un lielāka, sedimentācijas konstante 13S un augstāka.
Sintezē B1-limfocīti, plazmas šūnas un, iespējams, B1-limfocīti gļotādu un vi-
dalīties savos noslēpumos. Ražošanas apjoms var sasniegt 5 g uz dobu. SIgA kopums ir svarīgs lielākajam skaitam organismā – daži no tiem pārsniedz kopējo IgM un IgG daudzumu. Ir asinis sirovats s IgA nav parādīts.
SIgA molekulas ceturtdaļas struktūras veidošanās ir saistīta ar translokāciju caur epitēlija klitīnu. Uz epitēlija klitīna bazālās un sānu virsmas receptors tiek pārnests uz polimēra Ig molekulas (JR) J-lance. Tuvojoties receptoram, šūna endocitizē IgA, parādoties pūslīšiem, tas tiek pārnests uz epitēlija šūnas apikālo virsmu, de JR tiek pakļauts enzīmu šķelšanai. Rezultātā IgA izdalās orgāna lūmena gļotu sekrēcijā jau sekrēcijas formā, jo fragments, kas ir pievienots Ig molekulai, JR kļūst par S-lance.
IgA sekrēcijas forma ir galvenais faktors gļotādas un elpceļu gļotādas gļotādas un sehostatiskās sistēmas specifiskajā humorālajā piena dziedzeru imunitātē. Zavdyaki S-lanceus vin izturīgs pret diproteāzēm. sIgA neaktivizē komplementu, bet efektīvi saistās ar antigēniem, neitralizē abus un mikrobu atkārtotu adhēziju uz epitēlija šūnām.